

The requirement of membrane lymphotoxin for the presence of dendritic cells in lymphoid tissues
- PMID: 10477548
- PMCID: PMC2195624
- DOI: 10.1084/jem.190.5.629
The requirement of membrane lymphotoxin for the presence of dendritic cells in lymphoid tissues
Abstract
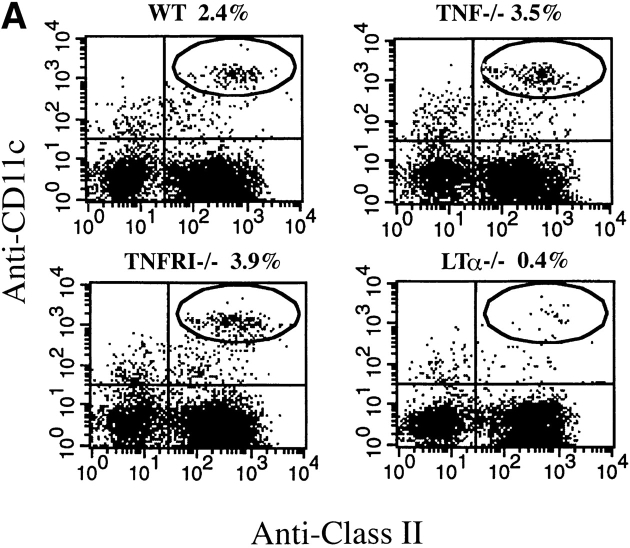
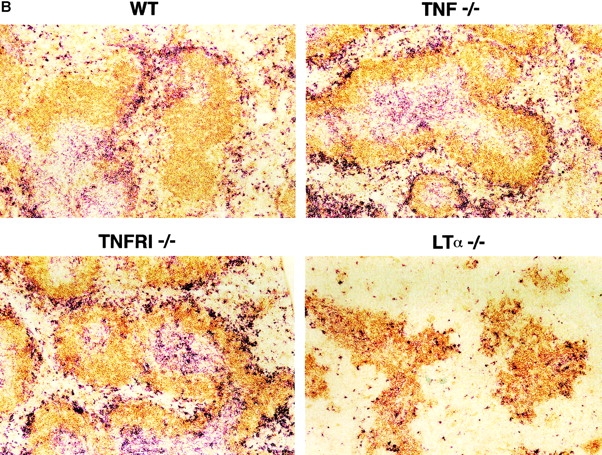
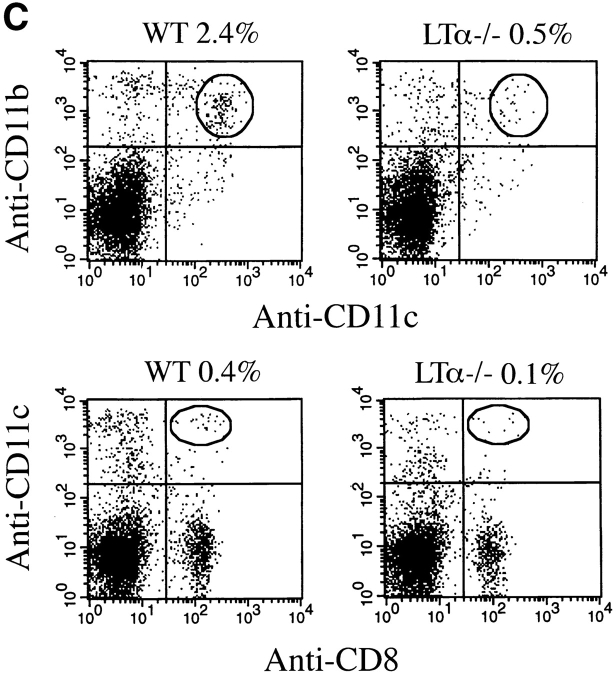
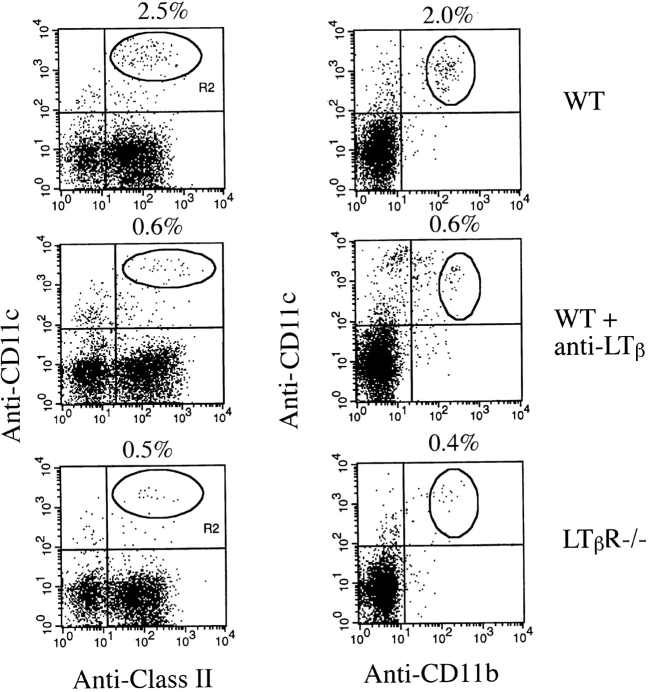
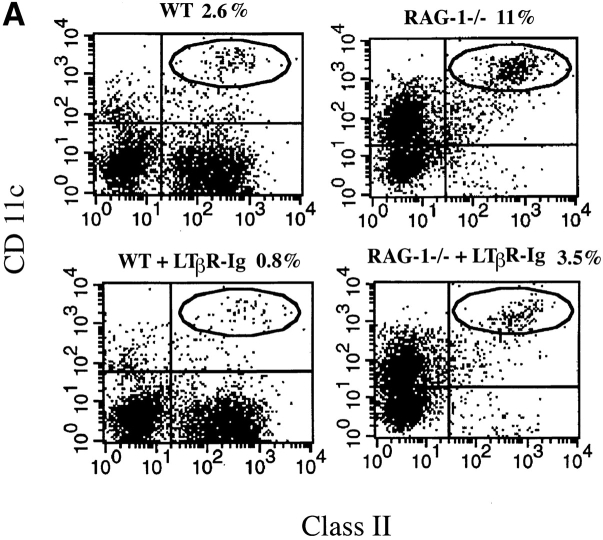
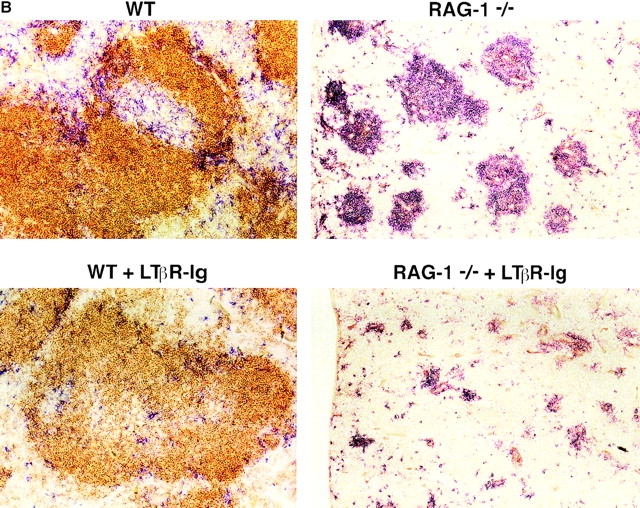
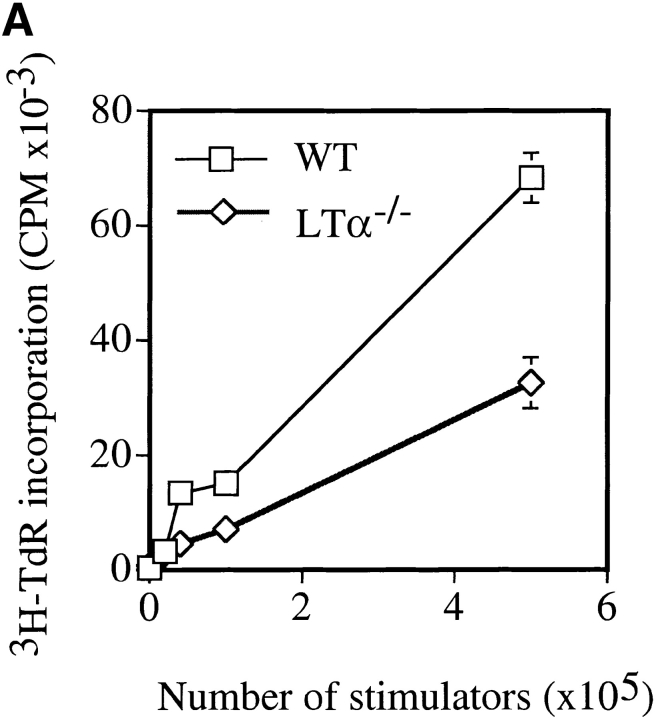
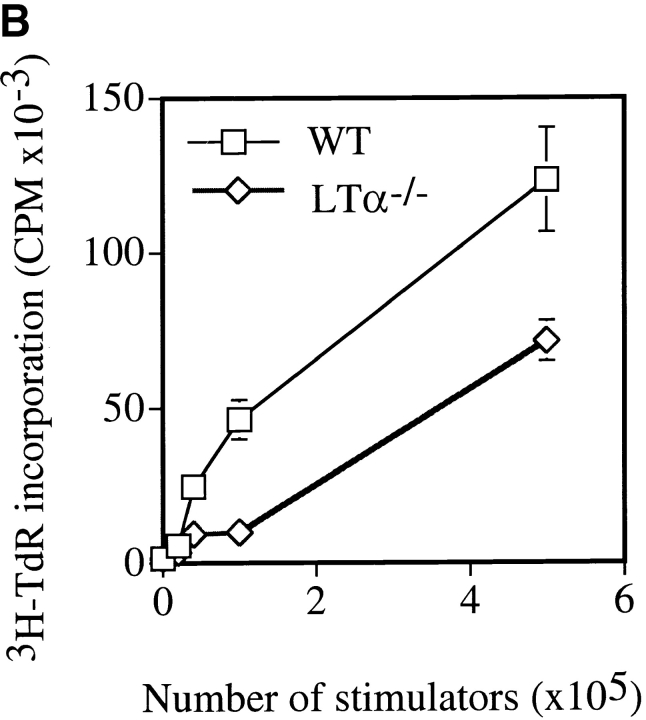
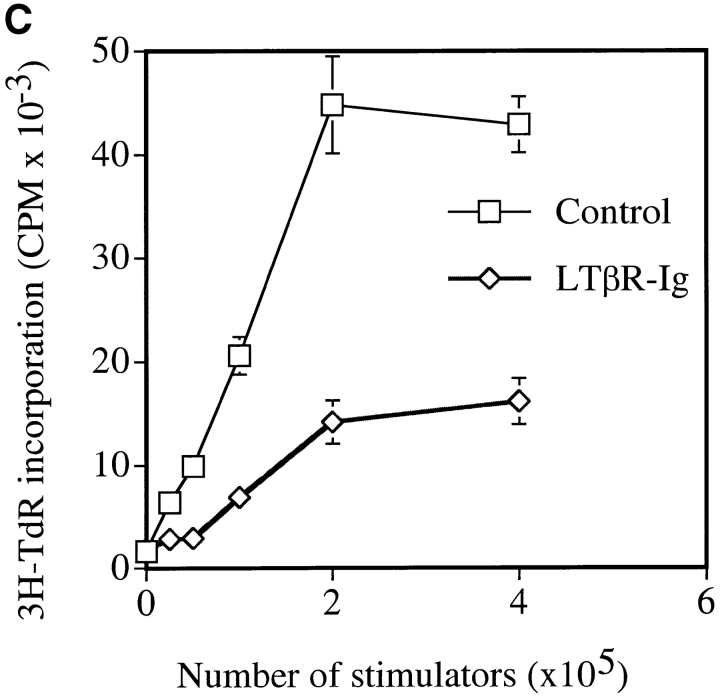
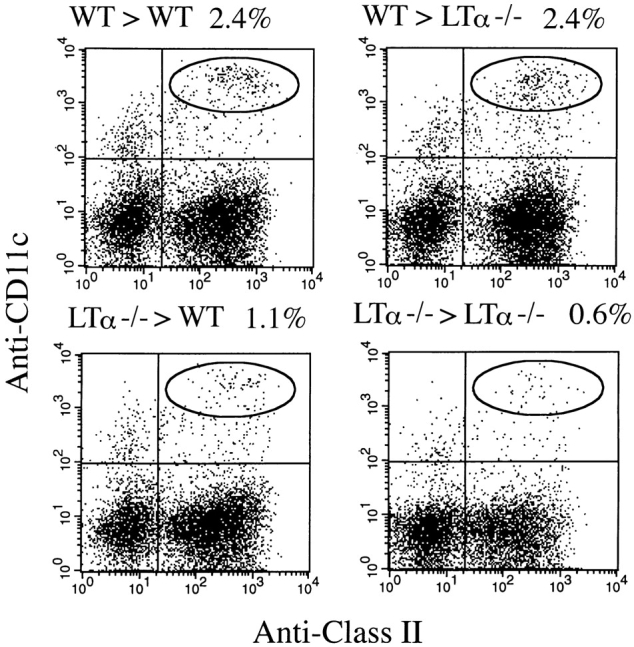
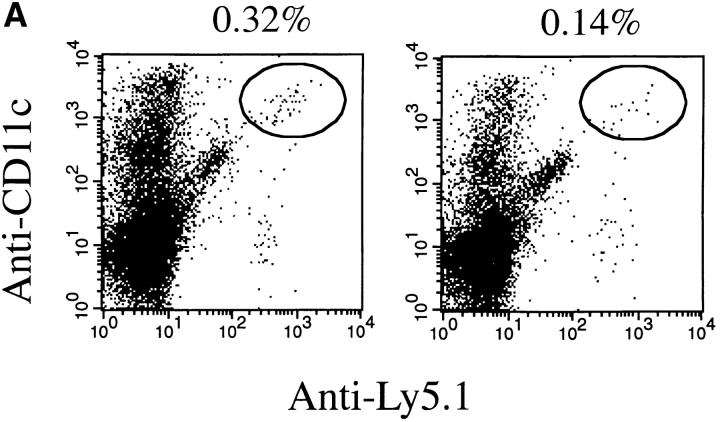
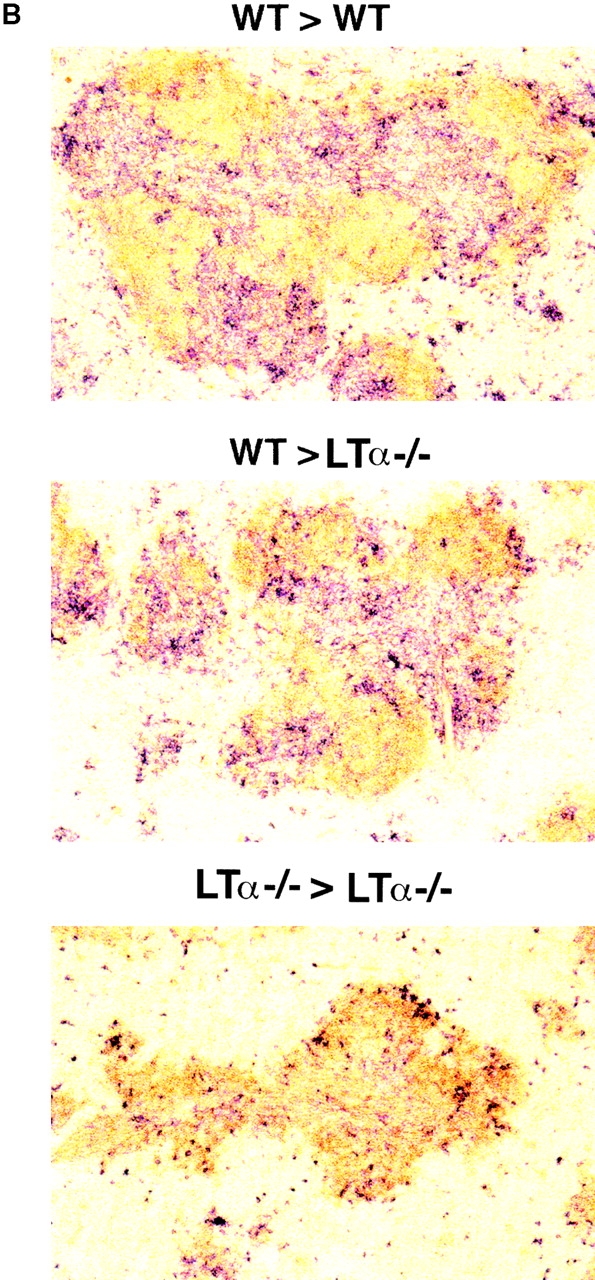
Although several cytokines, including tumor necrosis factor (TNF), can promote the growth of dendritic cells (DCs) in vitro, the cytokines that naturally regulate DC development and function in vivo have not been well defined. Here, we report that membrane lymphotoxin (LT), instead of TNF, regulates the migration of DCs in the spleen. LTalpha(-/-) mice, lacking membrane LTalpha/beta and LTalpha(3), show markedly reduced numbers of DCs in the spleen. Unlike wild-type mice and TNF(-/-) mice that have densely clustered DCs in the T cell zone and around the marginal zone, splenic DCs in LTalpha(-/-) mice are randomly distributed. The reduced number of DCs in lymphoid tissues of LTalpha(-/-) mice is associated with an increased number of DCs in nonlymphoid tissues. The number of splenic DCs in LTalpha(-/-) mice is restored when additional LT-expressing cells are provided. Blocking membrane LTalpha/beta in wild-type mice markedly diminishes the accumulation of DCs in lymphoid tissues. These data suggest that membrane LT is an essential ligand for the presence of DCs in the spleen. Mice deficient in TNF receptor, which is the receptor for both soluble LTalpha(3) and TNF-alpha(3) trimers, have normal numbers of DCs. However, LTbetaR(-/-) mice show reduced numbers of DCs, similar to the mice lacking membrane LT alpha/beta. Taken together, these results support the notion that the signaling via LTbetaR by membrane LTalpha/beta is required for the presence of DCs in lymphoid tissues.
Figures
References
-
- Ware C.F., VanArsdale T.L., Crowe P.D., Browning J.L. The ligands and receptors of the lymphotoxin system. Curr. Top. Microbiol. Immunol. 1995;198:175–218. - PubMed
-
- Fu Y.-X., Chaplin D.D. Development and maturation of secondary lymphoid tissues. Annu. Rev. Immunol. 1999;17:399–433. - PubMed
-
- Browning J.L., Ngam E.A., Lawton P., DeMarinis J., Tizard R., Chow E.P., Hession C., O'Brine G.B., Foley S.F., Ware C.F. Lymphotoxin beta, a novel member of the TNF family that forms a heteromeric complex with lymphotoxin on the cell surface. Cell. 1993;72:847–856. - PubMed
-
- Browning J.L., Sizing I.D., Lawton P., Bourdon P.R., Rennert P.D., Majeau G.R., Ambrose C.M., Hession C., Miatkowski K., Griffiths D.A. Characterization of lymphotoxin-alpha-beta complexes on the surface of mouse lymphocytes. J. Immunol. 1997;159:3288–3298. - PubMed
-
- Crowe P.D., VanArsdale T.L., Walter B.N., Ware C.F., Hession C., Ehrenfels B., Browning J.L., Din W.S., Goodwin R.G., Smith C.A. A lymphotoxin-beta-specific receptor. Science. 1994;264:707–710. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
