

Translational effects of mutations and polymorphisms in a repressive upstream open reading frame of the human cytomegalovirus UL4 gene
- PMID: 10482583
- PMCID: PMC112850
- DOI: 10.1128/JVI.73.10.8330-8337.1999
Translational effects of mutations and polymorphisms in a repressive upstream open reading frame of the human cytomegalovirus UL4 gene
Abstract
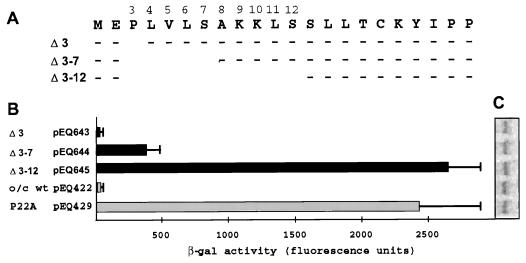
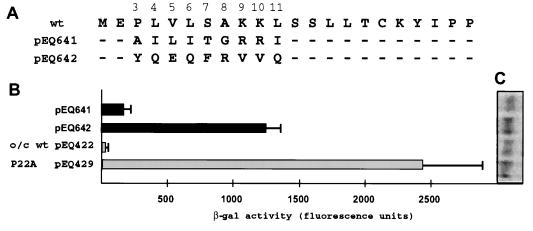
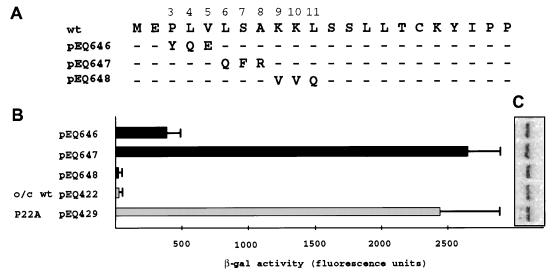
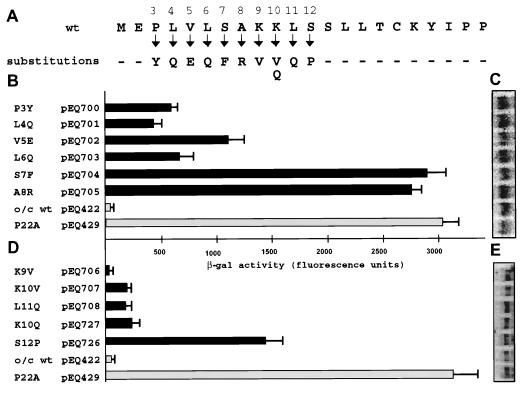
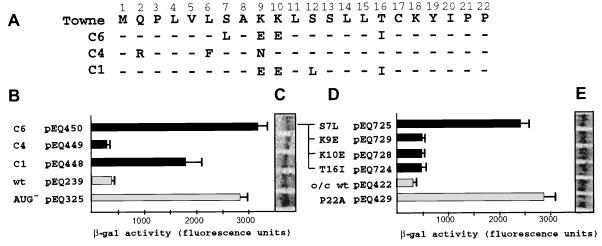
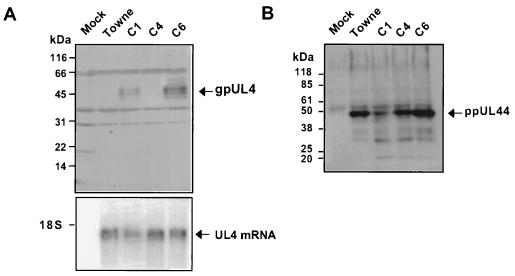
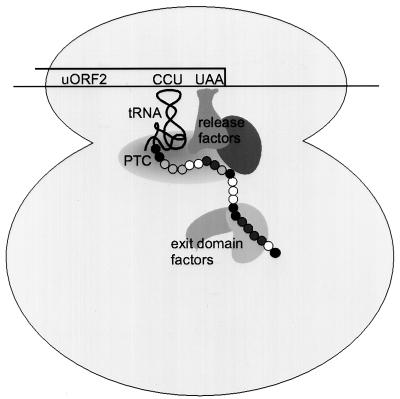
The human cytomegalovirus (HCMV) gpUL4 mRNA contains a 22-codon upstream open reading frame (uORF2), the peptide product of which represses downstream translation by blocking translation termination at its own stop codon and by causing ribosomes to stall on the mRNA. A distinctive feature of this unusual mechanism is its strict dependence on the uORF2 peptide sequence. To delineate sequence elements that function in the inhibitory mechanism, deletions and missense mutations affecting the previously uncharacterized amino-terminal region of uORF2 were analyzed in transient-transfection and infection assays. These experiments identified multiple codons in this region that are necessary for inhibition of downstream translation by uORF2 and, in conjunction with previous results, demonstrated that amino acids dispersed throughout the uORF2 peptide participate in the repressive mechanism. In contrast to the highly conserved carboxy terminus, the amino-terminal portion of the uORF2 peptide is polymorphic. A survey of uORF2 sequences in HCMV clinical isolates revealed that although most have uORF2 sequences that are predicted to retain the uORF2 inhibitory activity, approximately 15% contain polymorphisms at codons that are essential for full inhibition by uORF2. Consistent with predictions based on analyses of engineered mutations, two viral isolates with uORF2 sequences that do not inhibit downstream translation in transfection assays expressed much more gpUL4 protein but similar levels of UL4 mRNA compared to the levels produced by the prototypic laboratory strain HCMV (Towne) and another clinical isolate with an inhibitory variant uORF2. These results demonstrate that uORF2 is polymorphic in sequence and repressive activity and suggest that the uORF2 regulatory mechanism, although prevalent among natural HCMV isolates, is not absolutely essential for viral replication.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
