

Antisense repression of hexokinase 1 leads to an overaccumulation of starch in leaves of transgenic potato plants but not to significant changes in tuber carbohydrate metabolism
- PMID: 10482667
- PMCID: PMC59360
- DOI: 10.1104/pp.121.1.123
Antisense repression of hexokinase 1 leads to an overaccumulation of starch in leaves of transgenic potato plants but not to significant changes in tuber carbohydrate metabolism
Abstract
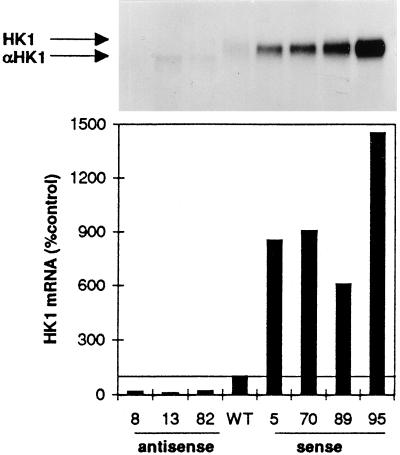
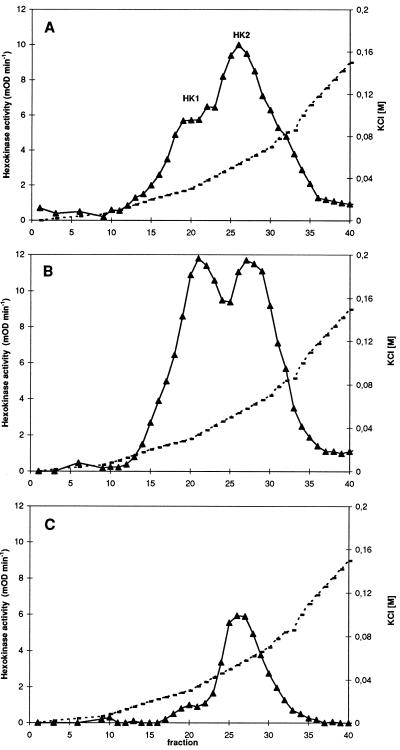
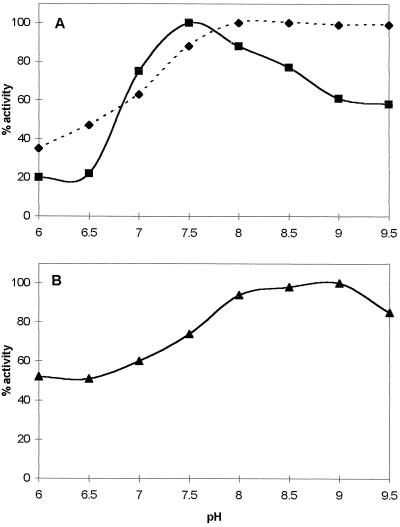
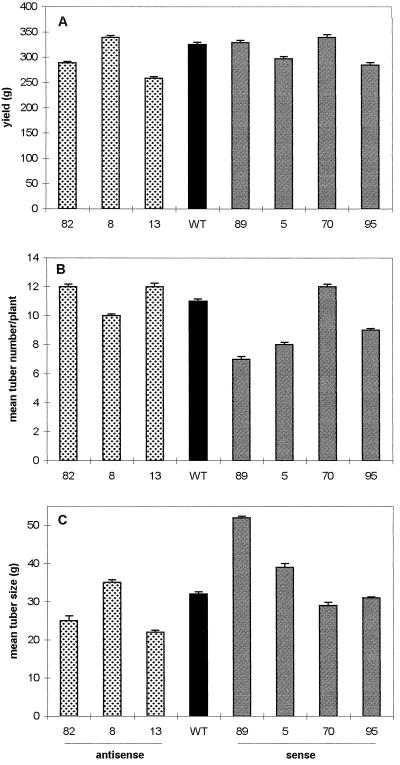
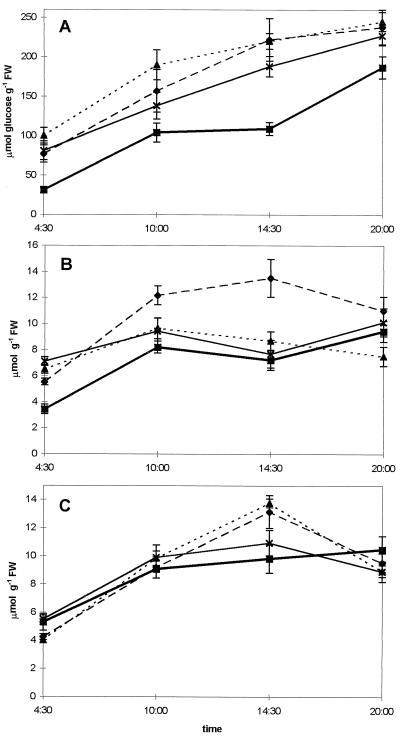
Potato (Solanum tuberosum L.) plants transformed with sense and antisense constructs of a cDNA encoding the potato hexokinase 1 (StHK1) exhibited altered enzyme activities and expression of StHK1 mRNA. Measurements of the maximum catalytic activity of hexokinase revealed a 22-fold variation in leaves (from 22% of the wild-type activity in antisense transformants to 485% activity in sense transformants) and a 7-fold variation in developing tubers (from 32% of the wild-type activity in antisense transformants to 222% activity in sense transformants). Despite the wide range of hexokinase activities, no change was found in the fresh weight yield, starch, sugar, or metabolite levels of transgenic tubers. However, there was a 3-fold increase in the starch content of leaves from the antisense transformants after the dark period. Starch accumulation at the end of the night period was correlated with a 2-fold increase of glucose and a decrease of sucrose content. These results provide strong support for the hypothesis that glucose is a primary product of transitory starch degradation and is the sugar that is exported to the cytosol at night to support sucrose biosynthesis.
Figures
References
-
- Baldus B, Kelly GJ, Latzko E. Hexokinases of spinach leaves. Phytochemistry. 1981;20:1811–1814.
-
- Bell GI, Pilkis SJ, Weber IT, Polonsky KS. Glucokinase mutations, insulin secretion, and diabetes mellitus. Annu Rev Physiol. 1996;58:171–186. - PubMed
-
- Burrell MM, Mooney PJ, Blundy M, Carter D, Wilson F, Green J, Blundy KS. Genetic manipulation of 6-phosphofructo-kinase in potato tubers. Planta. 1994;194:95–101.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
