

Pregnancy reduces brain sigma receptor function
- PMID: 10482906
- PMCID: PMC1566170
- DOI: 10.1038/sj.bjp.0702724
Pregnancy reduces brain sigma receptor function
Abstract
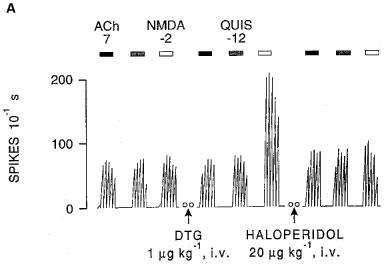
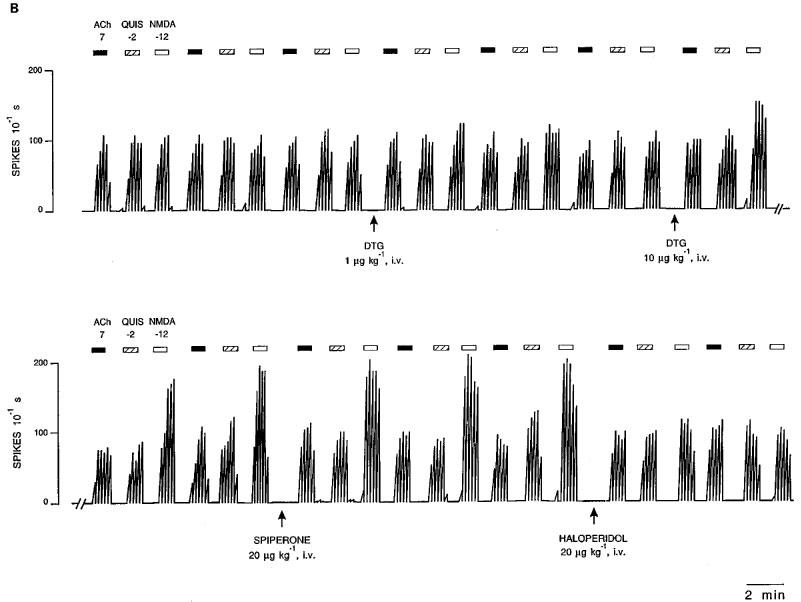
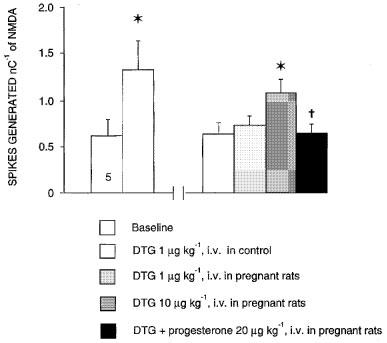
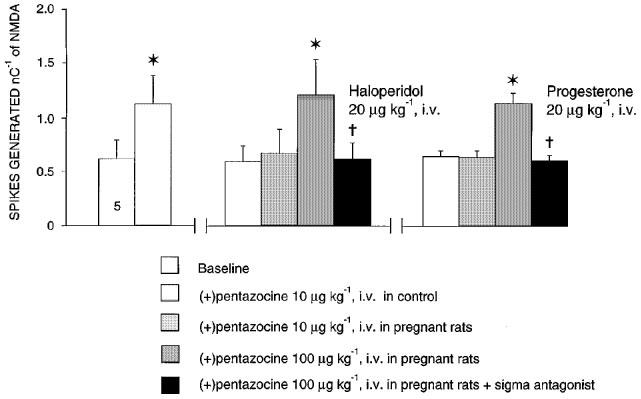
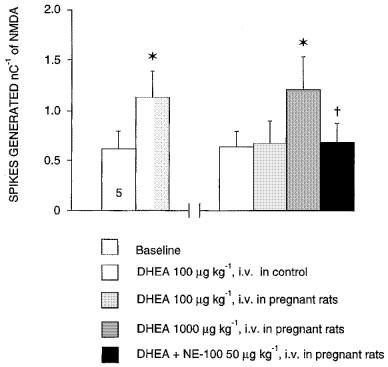
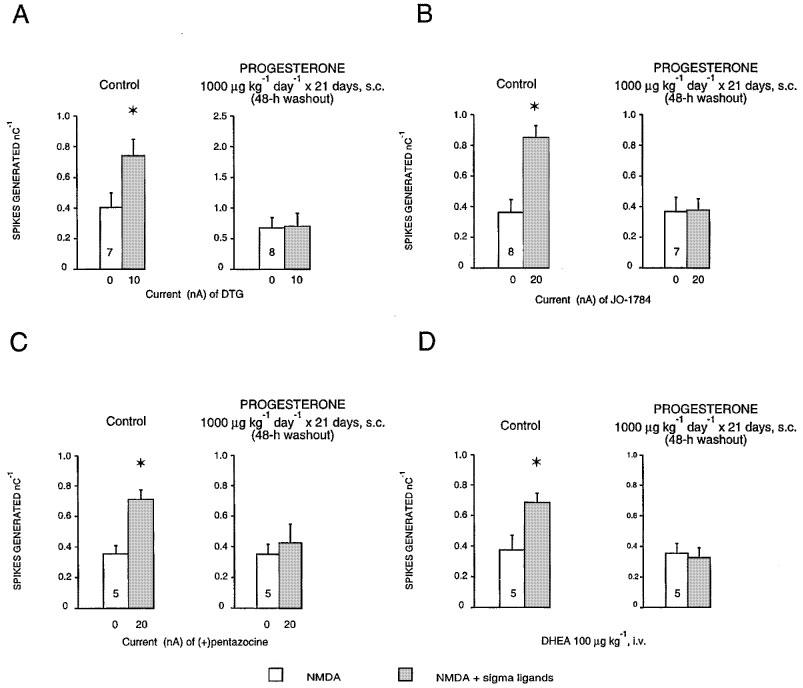
1. Sigma (sigma) receptors have recently been cloned, though their endogenous ligand(s) remain unidentified. However, some neuroactive steroids, such as progesterone, have a high affinity for these receptors. Some sigma ligands, such as DTG, (+)-pentazocine and DHEA, act as sigma 'agonists' by potentiating the neuronal response to NMDA. Others, such as haloperidol, NE-100 and progesterone, act as sigma 'antagonists' by reversing the potentiations induced by sigma 'agonists'. 2. We compared the effects of sigma 'agonists' in four series of female rats: in controls, at day 18 of pregnancy, at day 5 post-partum, and in ovariectomized rats following a 3-week treatment with a high dose of progesterone. 3. In pregnant rats and following a 3-week treatment with progesterone, 10 fold higher doses of DTG, (+)-pentazocine and DHEA were required to elicit a selective potentiation of the NMDA response comparable to that obtained in control females. Conversely, at day 5 post-partum and following the 3-week treatment with a progesterone and after a 5-day washout, the potentiation of the NMDA response induced by the sigma 'agonist' DTG was greater than in control females. 4. The present data suggest that endogenous progesterone acts as an 'antagonist' at sigma receptors. The resulting changes in the function of sigma receptors during pregnancy and post-partum may be implicated in emotional phenomena occurring during these periods.
Figures


References
-
- BACKSTROM T., BIXO M., HAMMARBACK S. Ovarian steroid hormones. Effects on mood, behaviour and brain excitability. Acta Obstet. Gynecologica Scand. 1985;130:19–24. - PubMed
-
- BERGERON R., DE MONTIGNY C., DEBONNEL G. Biphasic effects of sigma ligands on the neuronal response to N-methyl-D-aspartate. Naunyn Schmiedebergs Arch. Pharmacol. 1995;351:252–260. - PubMed
-
- CHAKI S., TANAKA M., MURAMATSU M., OTOMO S. NE-100, a novel potent σ ligand, preferentially binds to σ1 binding sites in guinea pig brain. Eur. J. Pharmacol. 1994;251:R1–R2. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
