

O(2) deprivation inhibits Ca(2+)-activated K(+) channels via cytosolic factors in mice neocortical neurons
- PMID: 10487772
- PMCID: PMC408544
- DOI: 10.1172/JCI7291
O(2) deprivation inhibits Ca(2+)-activated K(+) channels via cytosolic factors in mice neocortical neurons
Abstract
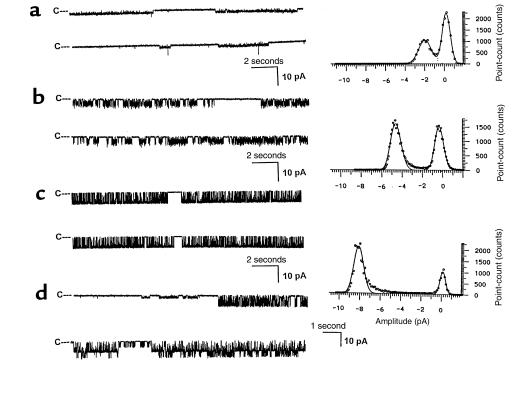
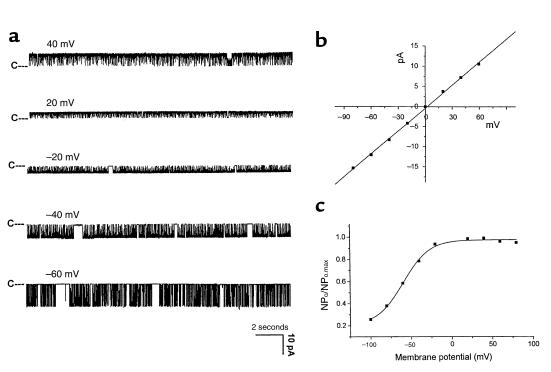
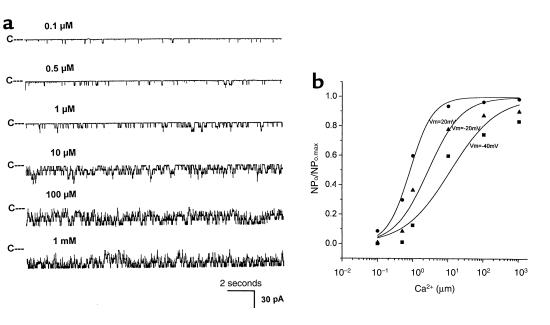
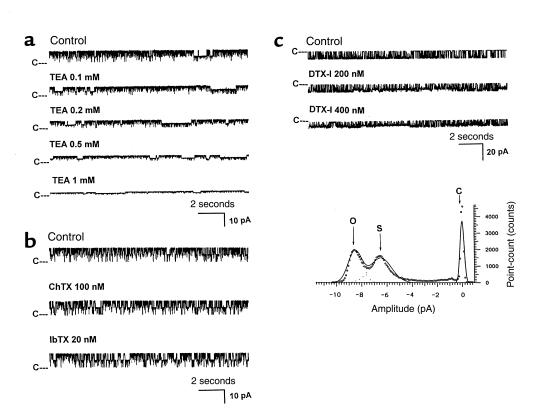
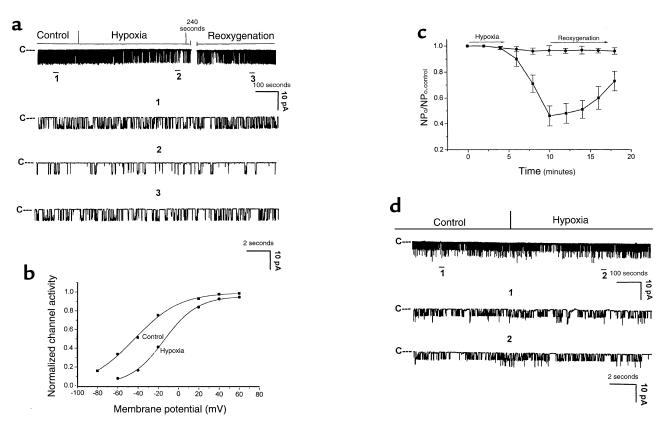
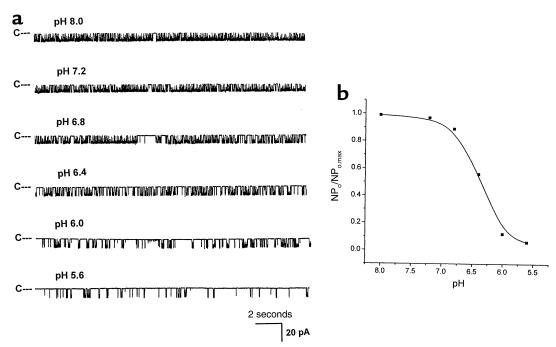
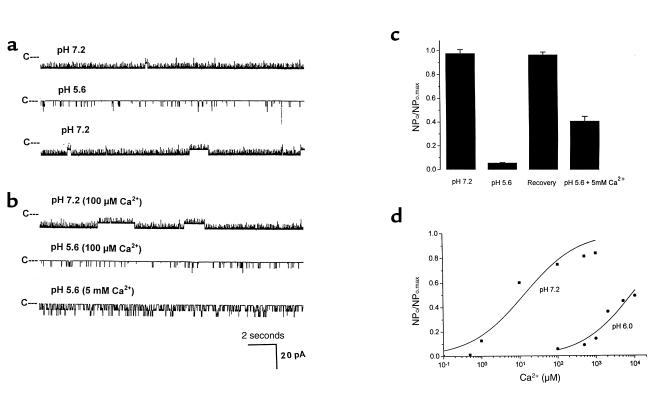
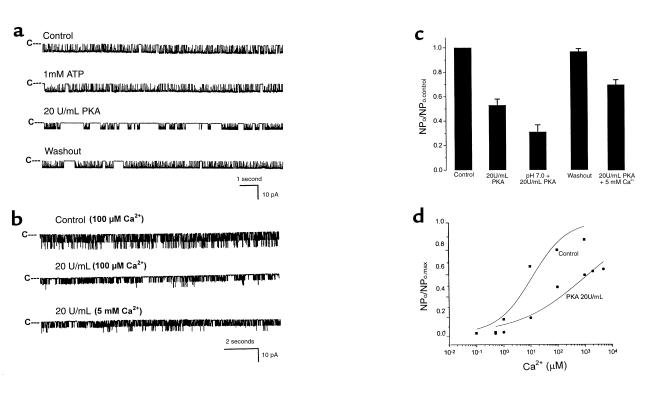
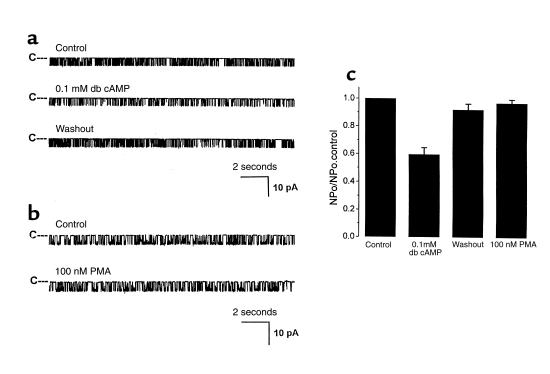
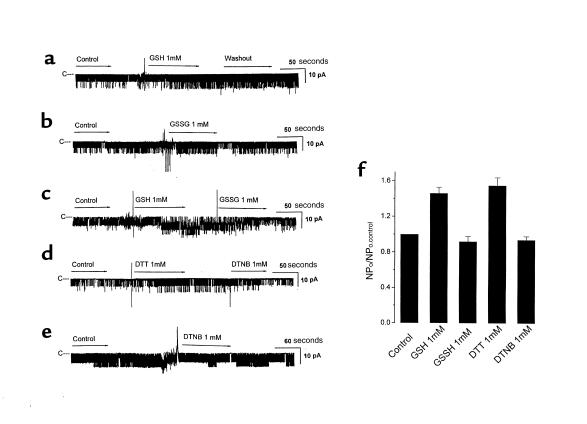
O(2) deprivation induces membrane depolarization in mammalian central neurons. It is possible that this anoxia-induced depolarization is partly mediated by an inhibition of K(+) channels. We therefore performed experiments using patch-clamp techniques and dissociated neurons from mice neocortex. Three types of K(+) channels were observed in both cell-attached and inside-out configurations, but only one of them was sensitive to lack of O(2). This O(2)-sensitive K(+) channel was identified as a large-conductance Ca(2+)-activated K(+) channel (BK(Ca)), as it exhibited a large conductance of 210 pS under symmetrical K(+) (140 mM) conditions, a strong voltage-dependence of activation, and a marked sensitivity to Ca(2+). A low-O(2) medium (PO(2) = 10-20 mmHg) markedly inhibited this BK(Ca) channel open probability in a voltage-dependent manner in cell-attached patches, but not in inside-out patches, indicating that the effect of O(2) deprivation on BK(Ca) channels of mice neocortical neurons was mediated via cytosol-dependent processes. Lowering intracellular pH (pH(i)), or cytosolic addition of the catalytic subunit of a cAMP-dependent protein kinase A in the presence of Mg-ATP, caused a decrease in BK(Ca) channel activity by reducing the sensitivity of this channel to Ca(2+). In contrast, the reducing agents glutathione and DTT increased single BK(Ca) channel open probability without affecting unitary conductance. We suggest that in neocortical neurons, (a) BK(Ca) is modulated by O(2) deprivation via cytosolic factors and cytosol-dependent processes, and (b) the reduction in channel activity during hypoxia is likely due to reduced Ca(2+) sensitivity resulting from cytosolic alternations such as in pH(i) and phosphorylation. Because of their large conductance and prevalence in the neocortex, BK(Ca) channels may be considered as a target for pharmacological intervention in conditions of acute anoxia or ischemia.
Figures
References
-
- Haddad GG, Jiang C. O2 deprivation in the central nervous system: on mechanisms of neuronal response, different sensitivity and injury. Prog Neurobiol. 1993;40:277–318. - PubMed
-
- Haddad GG, Jiang C. O2-sensing mechanisms in excitable cells: role of plasma membrane K+ channel. Annu Rev Physiol. 1997;59:23–43. - PubMed
-
- Lopez-Barneo J, Lopez-Lopez JR, Urena J, Gonzalez C. Chemotransduction in the carotid body: K+ current modulated by PO2 in type I chemoreceptor cells. Science. 1988;241:580–582. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
