

TANK potentiates tumor necrosis factor receptor-associated factor-mediated c-Jun N-terminal kinase/stress-activated protein kinase activation through the germinal center kinase pathway
- PMID: 10490605
- PMCID: PMC84649
- DOI: 10.1128/MCB.19.10.6665
TANK potentiates tumor necrosis factor receptor-associated factor-mediated c-Jun N-terminal kinase/stress-activated protein kinase activation through the germinal center kinase pathway
Abstract
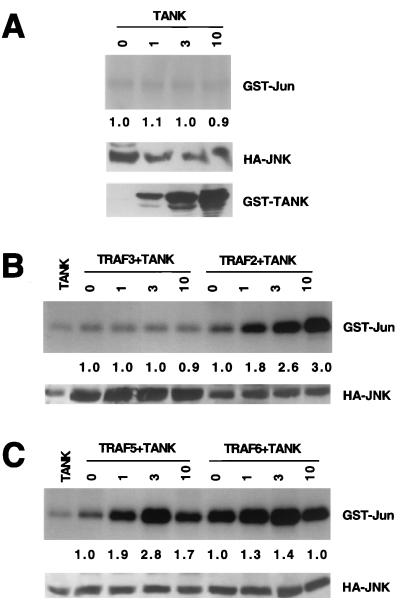
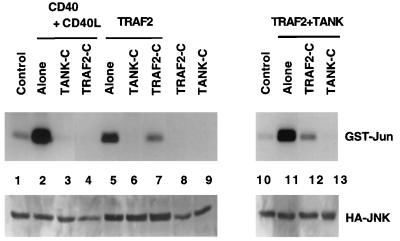
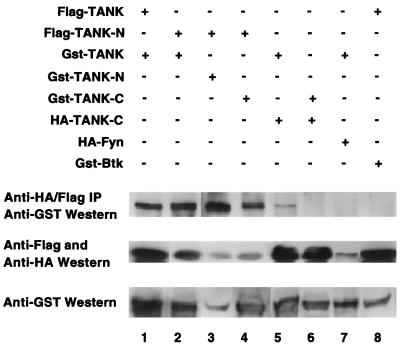
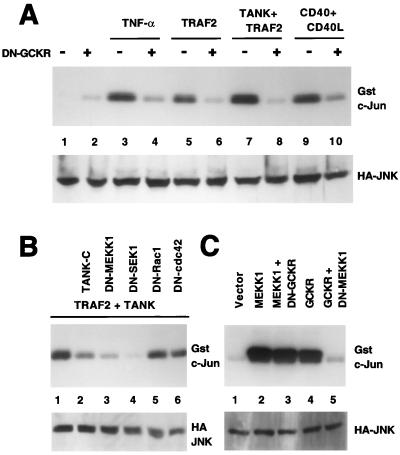
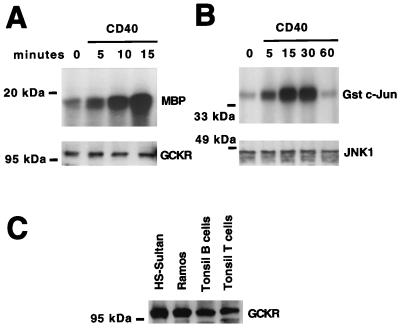
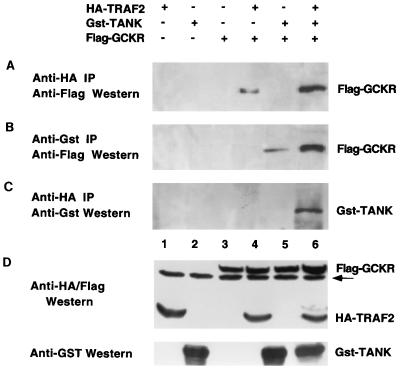
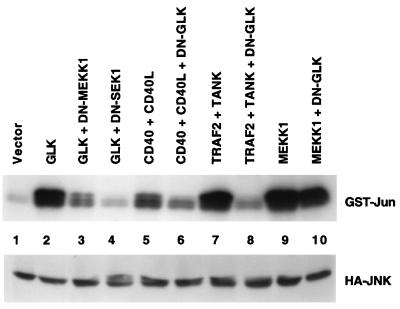
Tumor necrosis factor (TNF) receptor-associated factors (TRAFs) are mediators of many members of the TNF receptor superfamily and can activate both the nuclear factor kappaB (NF-kappaB) and stress-activated protein kinase (SAPK; also known as c-Jun N-terminal kinase) signal transduction pathways. We previously described the involvement of a TRAF-interacting molecule, TRAF-associated NF-kappaB activator (TANK), in TRAF2-mediated NF-kappaB activation. Here we show that TANK synergized with TRAF2, TRAF5, and TRAF6 but not with TRAF3 in SAPK activation. TRAF2 and TANK individually formed weak interactions with germinal center kinase (GCK)-related kinase (GCKR). However, when coexpressed, they formed a strong complex with GCKR, thereby providing a potential mechanism for TRAF and TANK synergy in GCKR-mediated SAPK activation, which is important in TNF family receptor signaling. Our results also suggest that TANK can form potential intermolecular as well as intramolecular interactions between its amino terminus and carboxyl terminus. This study suggests that TANK is a regulatory molecule controlling the threshold of NF-kappaB and SAPK activities in response to activation of TNF receptors. In addition, CD40 activated endogenous GCKR in primary B cells, implicating GCK family proteins in CD40-mediated B-cell functions.
Figures
Similar articles
-
Activation of stress-activated protein kinase/c-Jun N-terminal kinase, but not NF-kappaB, by the tumor necrosis factor (TNF) receptor 1 through a TNF receptor-associated factor 2- and germinal center kinase related-dependent pathway.J Biol Chem. 1997 Dec 19;272(51):32102-7. doi: 10.1074/jbc.272.51.32102. J Biol Chem. 1997. PMID: 9405407
-
Differential requirements for tumor necrosis factor receptor-associated factor family proteins in CD40-mediated induction of NF-kappaB and Jun N-terminal kinase activation.J Biol Chem. 1999 Aug 6;274(32):22414-22. doi: 10.1074/jbc.274.32.22414. J Biol Chem. 1999. PMID: 10428814
-
CD27, a member of the tumor necrosis factor receptor superfamily, activates NF-kappaB and stress-activated protein kinase/c-Jun N-terminal kinase via TRAF2, TRAF5, and NF-kappaB-inducing kinase.J Biol Chem. 1998 May 22;273(21):13353-8. doi: 10.1074/jbc.273.21.13353. J Biol Chem. 1998. PMID: 9582383
-
Tumor Necrosis Factor Receptor-Associated Factor Regulation of Nuclear Factor κB and Mitogen-Activated Protein Kinase Pathways.Front Immunol. 2018 Aug 9;9:1849. doi: 10.3389/fimmu.2018.01849. eCollection 2018. Front Immunol. 2018. PMID: 30140268 Free PMC article. Review.
-
Tumor necrosis factor receptor-associated factors (TRAFs).Oncogene. 2001 Oct 1;20(44):6482-91. doi: 10.1038/sj.onc.1204788. Oncogene. 2001. PMID: 11607847 Review.
Cited by
-
Toll-like receptors in sepsis-associated cytokine storm and their endogenous negative regulators as future immunomodulatory targets.Int Immunopharmacol. 2020 Dec;89(Pt B):107087. doi: 10.1016/j.intimp.2020.107087. Epub 2020 Oct 12. Int Immunopharmacol. 2020. PMID: 33075714 Free PMC article. Review.
-
Transcriptome and DNA Methylome Analysis in a Mouse Model of Diet-Induced Obesity Predicts Increased Risk of Colorectal Cancer.Cell Rep. 2018 Jan 16;22(3):624-637. doi: 10.1016/j.celrep.2017.12.071. Cell Rep. 2018. PMID: 29346762 Free PMC article.
-
Regulation of the PKCθ-NF-κB Axis in T Lymphocytes by the Tumor Necrosis Factor Receptor Family Member OX40.Front Immunol. 2012 May 28;3:133. doi: 10.3389/fimmu.2012.00133. eCollection 2012. Front Immunol. 2012. PMID: 22654884 Free PMC article.
-
TRAF family member-associated NF-kappa B activator (TANK) expression increases in injured sensory neurons and is transcriptionally regulated by Sox11.Neuroscience. 2013 Feb 12;231:28-37. doi: 10.1016/j.neuroscience.2012.11.034. Epub 2012 Nov 29. Neuroscience. 2013. PMID: 23201825 Free PMC article.
-
TRAF family member-associated NF-κB activator (TANK) induced by RANKL negatively regulates osteoclasts survival and function.Int J Biol Sci. 2012;8(10):1398-407. doi: 10.7150/ijbs.5079. Epub 2012 Nov 6. Int J Biol Sci. 2012. PMID: 23139637 Free PMC article.
References
-
- Anderson D, Maraskovsky E, Billingsley W, Dougall W, Tometsko M, Roux E, Teepe M, DuBose R, Cosman D, Galibert L. A homologue of the TNF receptor and its ligand enhance T-cell growth and dendritic-cell function. Nature. 1997;390:175–179. - PubMed
-
- Arch R H, Gehrich R W, Thompson C B. Tumor necrosis factor receptor-associated factors (TRAFs)—a family of adaptor proteins that regulates life and death. Genes Dev. 1998;12:2821–2830. - PubMed
-
- Bagrodia S, Derijard B, Davis R J, Cerione R A. Cdc42 and PAK-mediated signaling leads to Jun kinase and p38 mitogen-activated protein kinase activation. J Biol Chem. 1995;270:27995–27998. - PubMed
-
- Cao Z, Xiong J, Takeuchi M, Kurama T, Goeddel D V. TRAF6 is a signal transducer for interleukin-1. Nature. 1996;383:443–446. - PubMed
-
- Cheng G, Baltimore D. TANK, a co-inducer with TRAF2 of TNF- and CD40L-mediated NF-κB activation. Genes Dev. 1996;10:963–973. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous