

Targeted disruption of CDK4 delays cell cycle entry with enhanced p27(Kip1) activity
- PMID: 10490638
- PMCID: PMC84696
- DOI: 10.1128/MCB.19.10.7011
Targeted disruption of CDK4 delays cell cycle entry with enhanced p27(Kip1) activity
Abstract
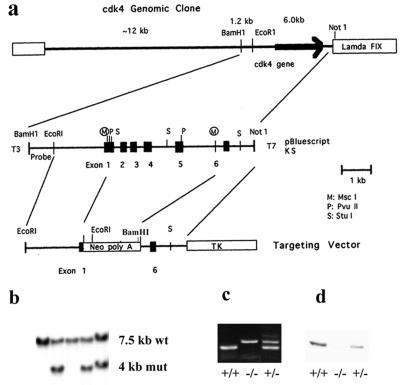
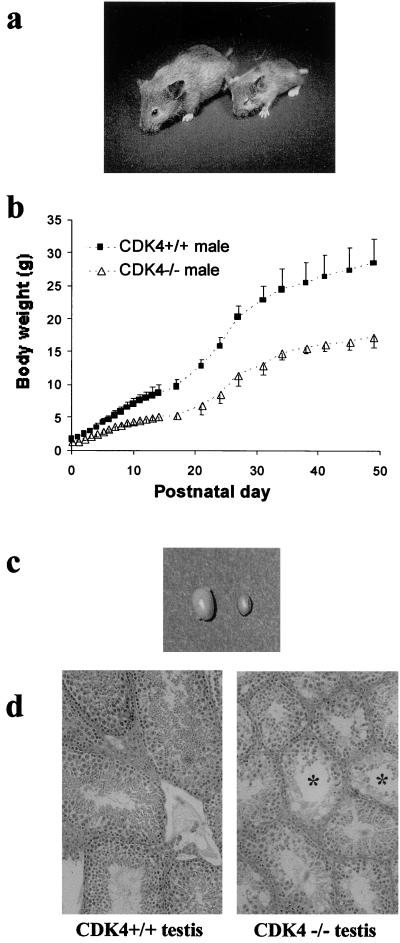
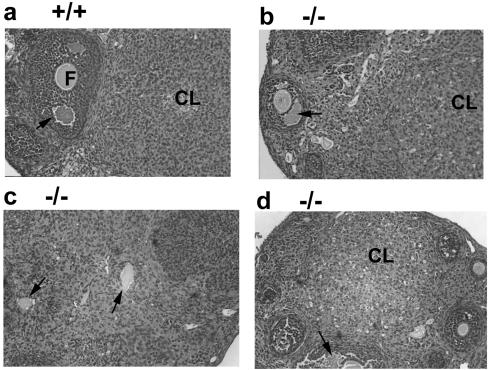
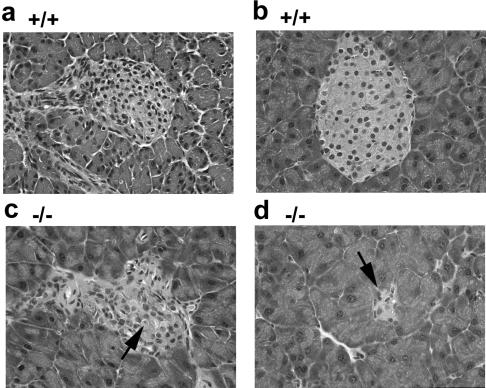
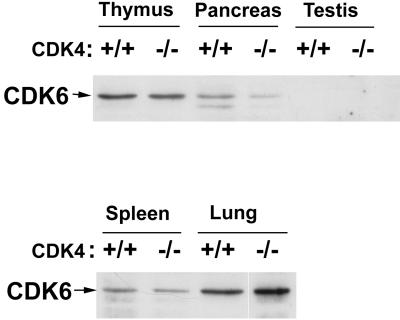
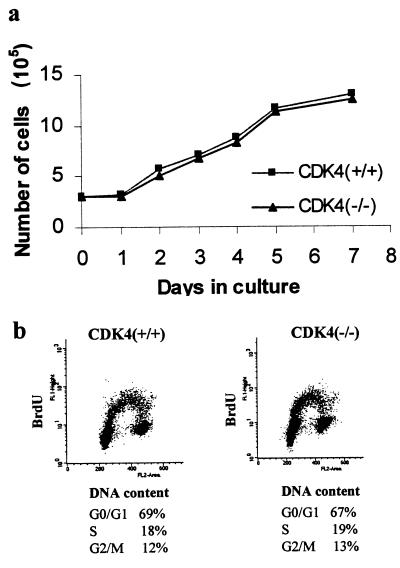
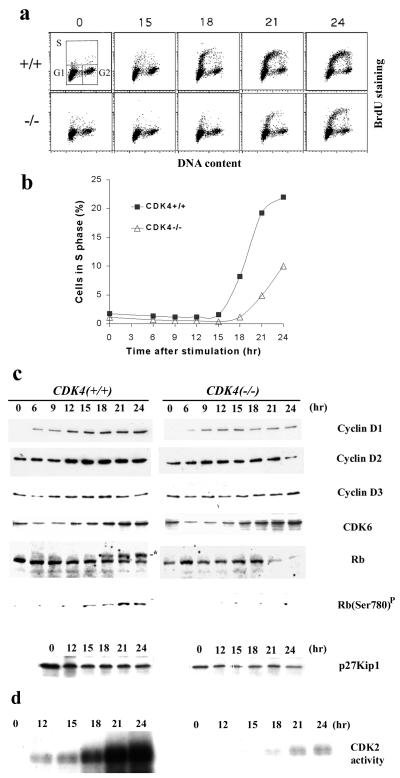
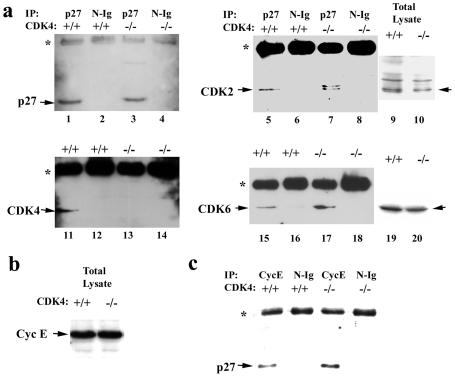
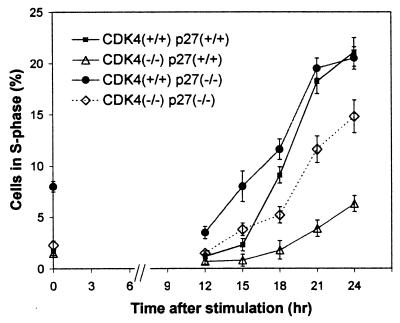
The mechanism by which cyclin-dependent kinase 4 (CDK4) regulates cell cycle progression is not entirely clear. Cyclin D/CDK4 appears to initiate phosphorylation of retinoblastoma protein (Rb) leading to inactivation of the S-phase-inhibitory action of Rb. However, cyclin D/CDK4 has been postulated to act in a noncatalytic manner to regulate the cyclin E/CDK2-inhibitory activity of p27(Kip1) by sequestration. In this study we investigated the roles of CDK4 in cell cycle regulation by targeted disruption of the mouse CDK4 gene. CDK4(-/-) mice survived embryogenesis and showed growth retardation and reproductive dysfunction associated with hypoplastic seminiferous tubules in the testis and perturbed corpus luteum formation in the ovary. These phenotypes appear to be opposite to those of p27-deficient mice such as gigantism and gonadal hyperplasia. A majority of CDK4(-/-) mice developed diabetes mellitus by 6 weeks, associated with degeneration of pancreatic islets. Fibroblasts from CDK4(-/-) mouse embryos proliferated similarly to wild-type embryonic fibroblasts under conditions that promote continuous growth. However, quiescent CDK4(-/-) fibroblasts exhibited a substantial ( approximately 6-h) delay in S-phase entry after serum stimulation. This cell cycle perturbation by CDK4 disruption was associated with increased binding of p27 to cyclin E/CDK2 and diminished activation of CDK2 accompanied by impaired Rb phosphorylation. Importantly, fibroblasts from CDK4(-/-) p27(-/-) embryos displayed partially restored kinetics of the G(0)-S transition, indicating the significance of the sequestration of p27 by CDK4. These results suggest that at least part of CDK4's participation in the rate-limiting mechanism for the G(0)-S transition consists of controlling p27 activity.
Figures
References
-
- Bates S, Bonetta L, MacAllan D, Parry D, Holder A, Dickson C, Peters G. CDK6 (PLSTIRE) and CDK4 (PSK-J3) are a distinct subset of the cyclin-dependent kinases that associate with cyclin D1. Oncogene. 1994;9:71–79. - PubMed
-
- Beijersbergen R L, Carlee L, Kerkhoven R M, Bernards R. Regulation of the retinoblastoma protein-related p107 by G1 cyclin complexes. Genes Dev. 1995;9:1340–1353. - PubMed
-
- Bignon Y J, Chen Y, Chang C Y, Riley D J, Windle J J, Mellon P L, Lee W H. Expression of a retinoblastoma transgene results in dwarf mice. Genes Dev. 1993;7:1654–1662. - PubMed
-
- Blain S W, Montalvo E, Massague J. Differential interaction of the cyclin-dependent kinase (Cdk) inhibitor p27Kip1 with cyclin A-Cdk2 and cyclin D2-Cdk4. J Biol Chem. 1997;272:25863–25872. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous