

Cellular and molecular mechanisms of glial scarring and progressive cavitation: in vivo and in vitro analysis of inflammation-induced secondary injury after CNS trauma
- PMID: 10493720
- PMCID: PMC6783021
- DOI: 10.1523/JNEUROSCI.19-19-08182.1999
Cellular and molecular mechanisms of glial scarring and progressive cavitation: in vivo and in vitro analysis of inflammation-induced secondary injury after CNS trauma
Abstract
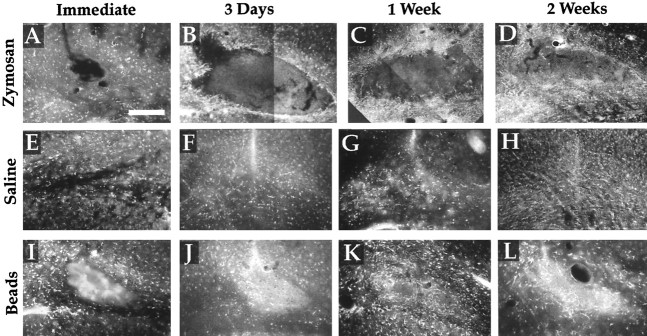
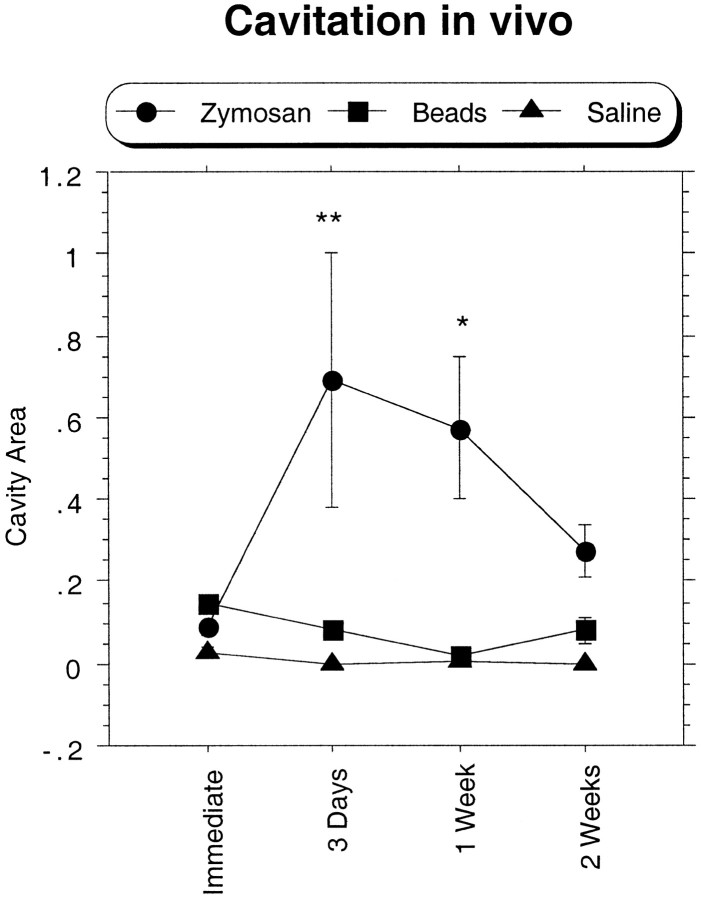
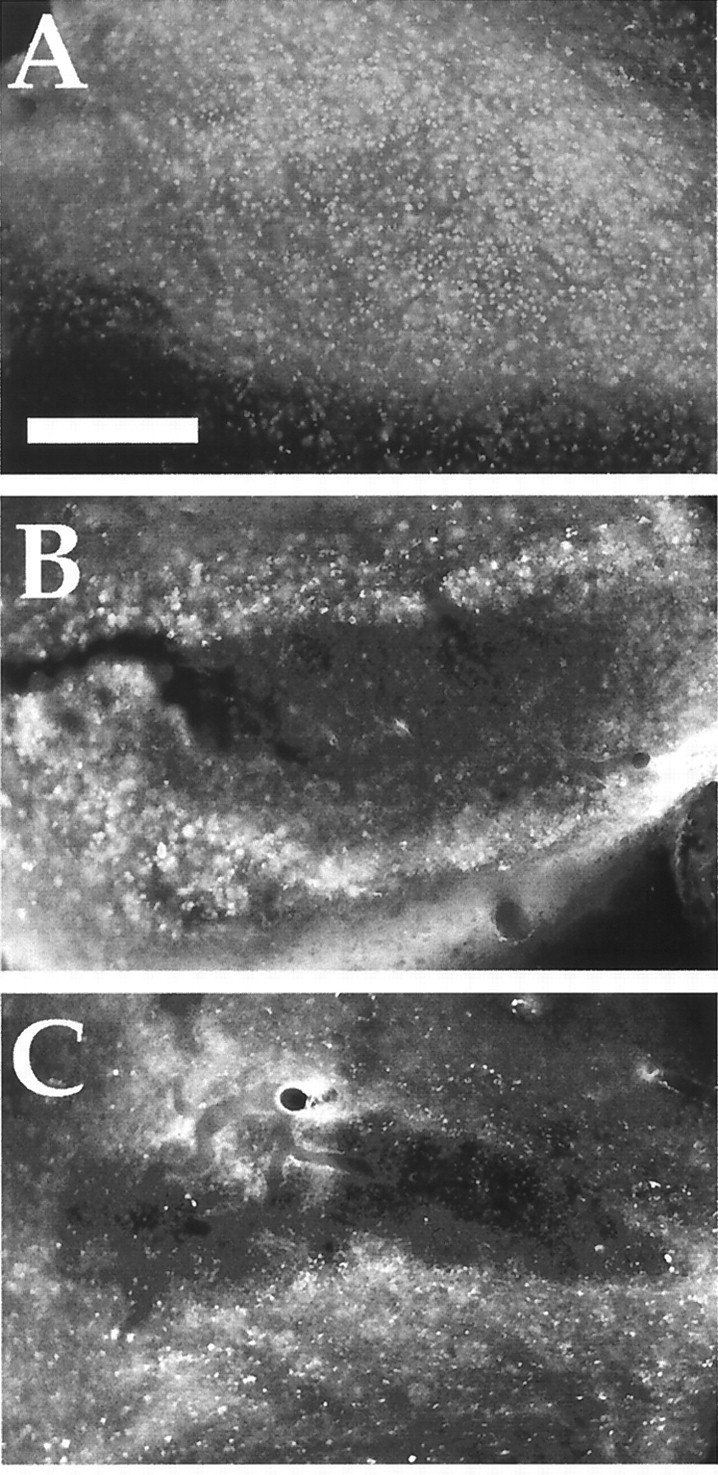
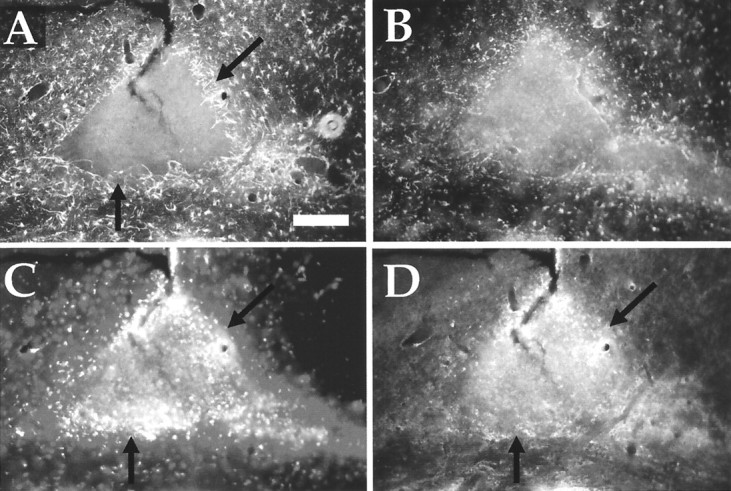
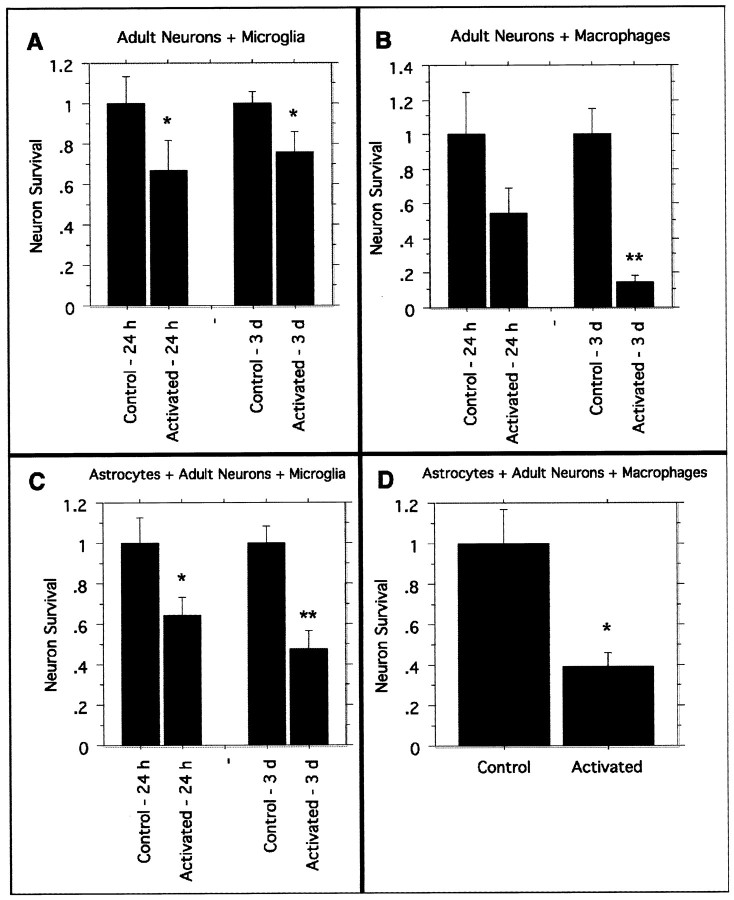
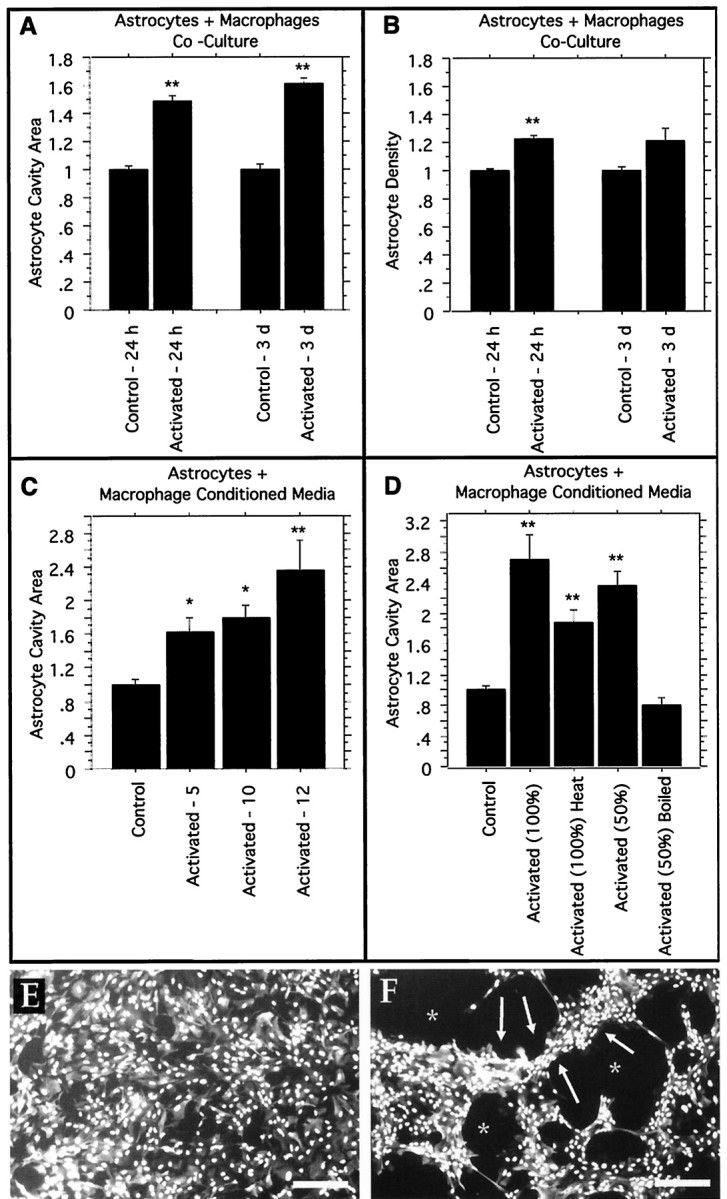
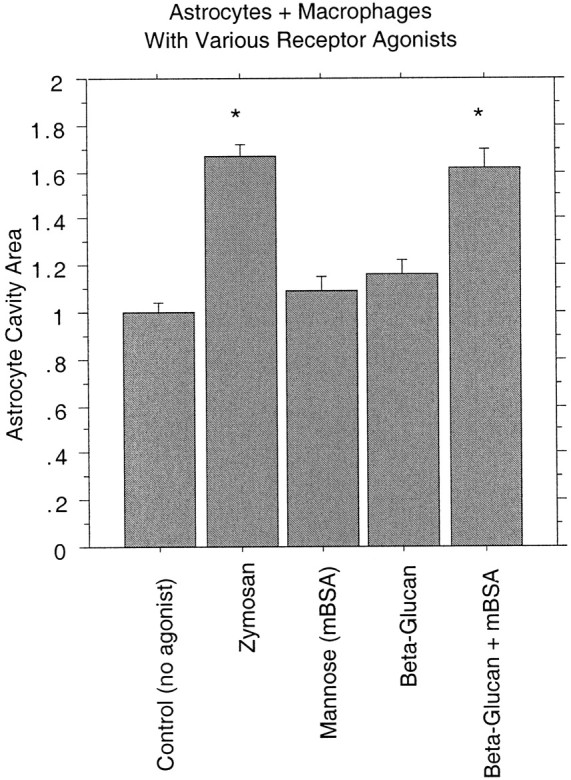
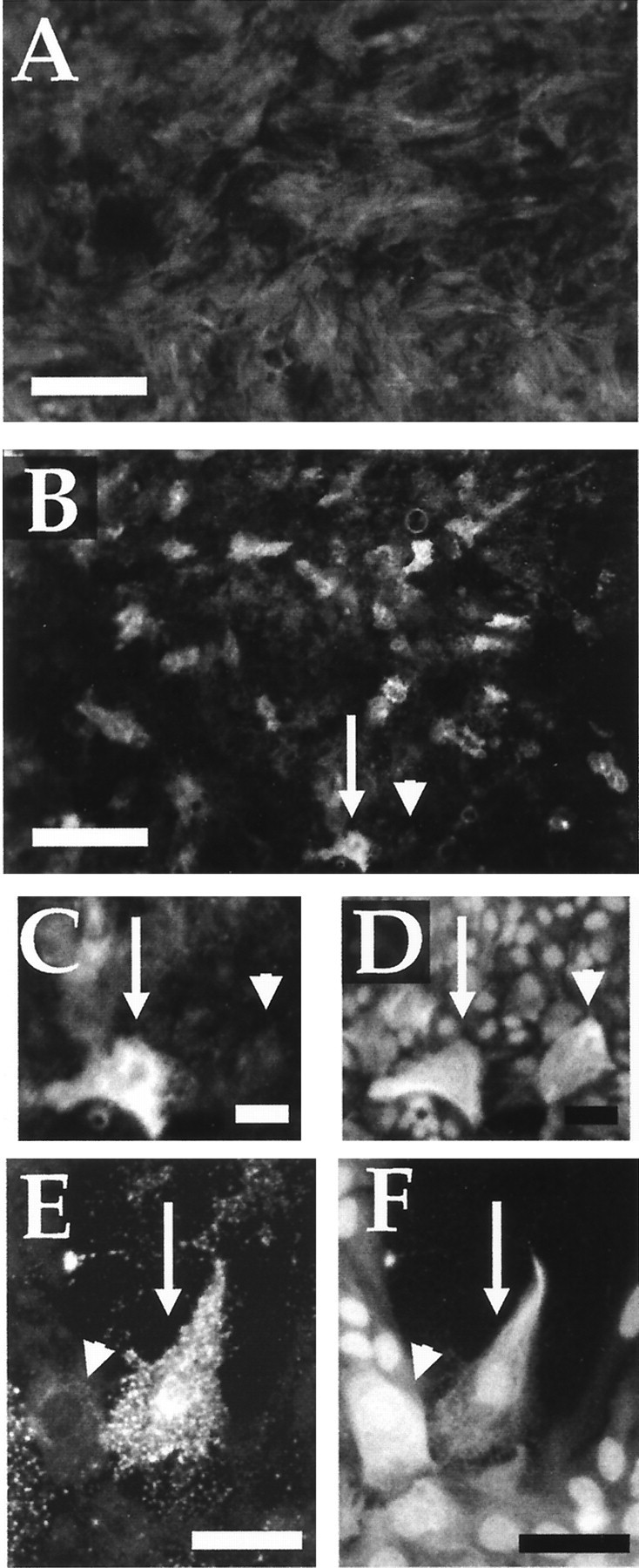
Post-traumatic cystic cavitation, in which the size and severity of a CNS injury progress from a small area of direct trauma to a greatly enlarged secondary injury surrounded by glial scar tissue, is a poorly understood complication of damage to the brain and spinal cord. Using minimally invasive techniques to avoid primary physical injury, this study demonstrates in vivo that inflammatory processes alone initiate a cascade of secondary tissue damage, progressive cavitation, and glial scarring in the CNS. An in vitro model allowed us to test the hypothesis that specific molecules that stimulate macrophage inflammatory activation are an important step in initiating secondary neuropathology. Time-lapse video analyses of inflammation-induced cavitation in our in vitro model revealed that this process occurs primarily via a previously undescribed cellular mechanism involving dramatic astrocyte morphological changes and rapid migration. The physical process of cavitation leads to astrocyte abandonment of neuronal processes, neurite stretching, and secondary injury. The macrophage mannose receptor and the complement receptor type 3 beta2-integrin are implicated in the cascade that induces cavity and scar formation. We also demonstrate that anti-inflammatory agents modulating transcription via the nuclear hormone receptor peroxisome proliferator-activated receptor-gamma may be therapeutic in preventing progressive cavitation by limiting inflammation and subsequent secondary damage after CNS injury.
Figures





References
-
- Alvarez-Buylla A, Temple S. Stem cells in the developing and adult nervous system. J Neurobiol. 1998;36:105–110. - PubMed
-
- Andersson PB, Perry VH, Gordon S. The acute inflammatory response to lipopolysaccharide in CNS parenchyma differs from that in other body tissues. Neuroscience. 1992;48:169–186. - PubMed
-
- Anthony DC, Bolton SJ, Fearn S, Perry VH. Age-related effects of interleukin-1 beta on polymorphonuclear neutrophil-dependent increases in blood-brain barrier permeability in rats. Brain. 1997;120:435–444. - PubMed
-
- Balentine JD. Pathology of experimental spinal cord trauma. I. The necrotic lesion as a function of vascular injury. Lab Invest. 1978;39:236–253. - PubMed
-
- Bartholdi D, Rubin BP, Schwab ME. VEGF mRNA induction correlates with changes in the vascular architecture upon spinal cord damage in the rat. Eur J Neurosci. 1997;9:2549–2560. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources