

Cholinergic induction of theta-frequency oscillations in hippocampal inhibitory interneurons and pacing of pyramidal cell firing
- PMID: 10493764
- PMCID: PMC6783040
- DOI: 10.1523/JNEUROSCI.19-19-08637.1999
Cholinergic induction of theta-frequency oscillations in hippocampal inhibitory interneurons and pacing of pyramidal cell firing
Abstract
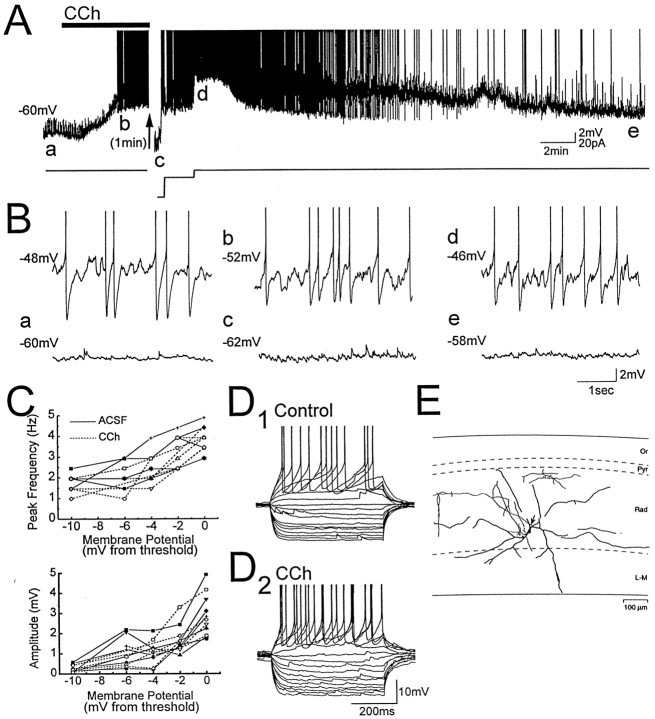
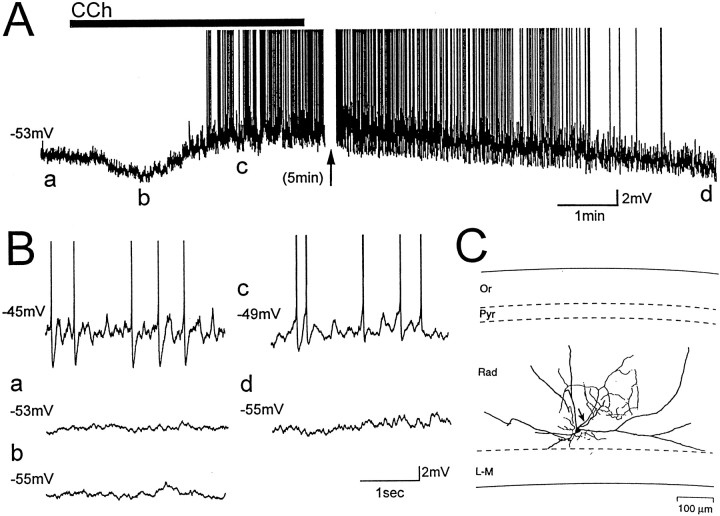
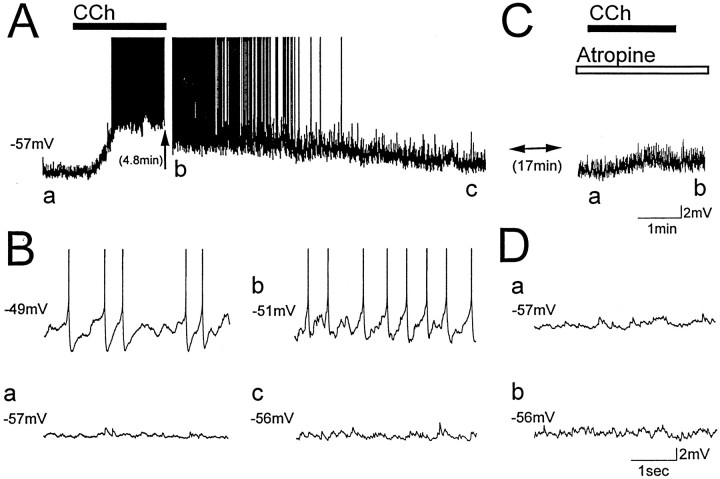
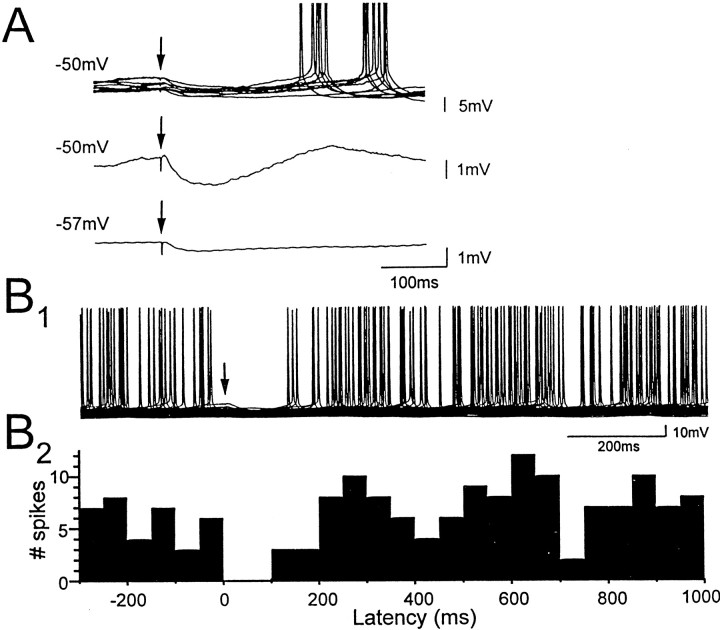
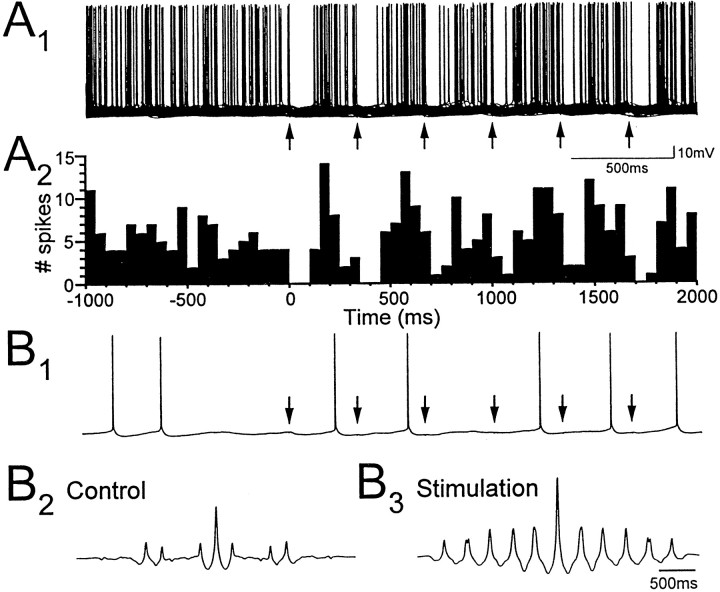
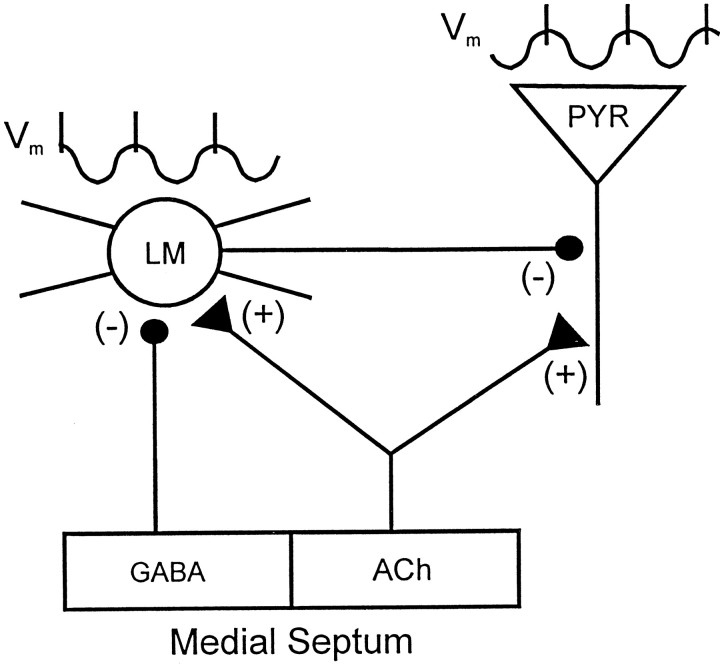
Cholinergic and GABAergic medial septal afferents contribute to hippocampal theta activity in part by actions on local interneurons. Interneurons near the border between stratum radiatum and stratum lacunosum-moleculare (LM) display intrinsic membrane potential oscillations at theta frequency when depolarized near threshold. First, whole-cell current-clamp recordings in rat hippocampal slices were used to examine effects of the cholinergic agonist carbachol on biocytin-labeled LM interneurons. At resting membrane potential, cells were depolarized by bath application of 25 microM carbachol, and the depolarization was sufficient to induce membrane potential oscillations (2.4 +/- 0.2 mV) that paced cell firing. Carbachol also depolarized LM interneurons in the presence of 6-cyano-7-nitroquinoxaline-2,3-dione, (+/-)-2-amino-5-phosphonopentanoic acid, and bicuculline, indicating that cholinergic depolarization of LM cells does not depend on ionotropic glutamate or GABA(A) synaptic transmission in local circuits. Atropine blocked the depolarization, indicating that muscarinic receptors were involved. Minimal stimulation applied to visually identified LM interneurons was then used to determine if spontaneous activity in CA1 pyramidal cells can be paced by rhythmic inhibition generated by LM cells at theta frequency. Inhibitory postsynaptic potentials evoked in pyramidal cells by single minimal stimulations were followed by rebound depolarizations and action potentials. When trains of minimal stimulation were delivered, membrane potential oscillations of depolarized pyramidal cells followed the stimulation frequency. Minimal stimulation led pyramidal cell firing with an average phase of 177 degrees. Thus, muscarinic induction of theta-frequency membrane potential oscillations in LM interneurons may contribute to the generation of rhythmic inhibition that paces intrinsically generated theta activity in CA1 pyramidal cells.
Figures
References
-
- Atzori M. Pyramidal cells and stratum lacunosum-moleculare interneurons in the CA1 hippocampal region share a GABAergic spontaneous input. Hippocampus. 1996;6:72–78. - PubMed
-
- Behrends JC, ten Bruggencate G. Cholinergic modulation of synaptic inhibition in the guinea pig hippocampus in vitro: excitation of GABAergic interneurons and inhibition of GABA-release. J Neurophysiol. 1993;69:626–629. - PubMed
-
- Bland BH, Colom LV. Extrinsic and intrinsic properties underlying oscillation and synchrony in limbic cortex. Prog Neurobiol. 1993;41:157–208. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous