

Pseudomonas aeruginosa induces type-III-secretion-mediated apoptosis of macrophages and epithelial cells
- PMID: 10496945
- PMCID: PMC96920
- DOI: 10.1128/IAI.67.10.5530-5537.1999
Pseudomonas aeruginosa induces type-III-secretion-mediated apoptosis of macrophages and epithelial cells
Abstract
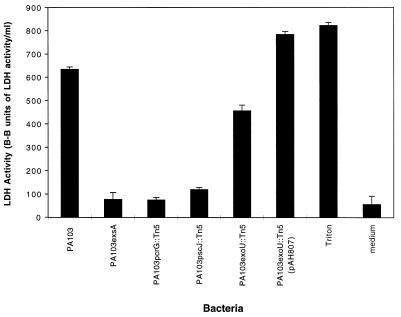
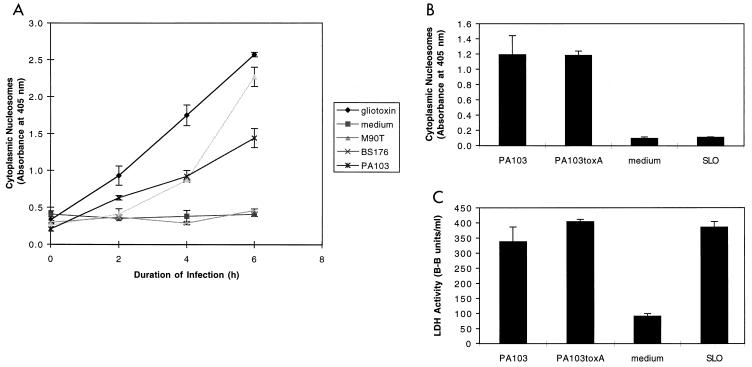
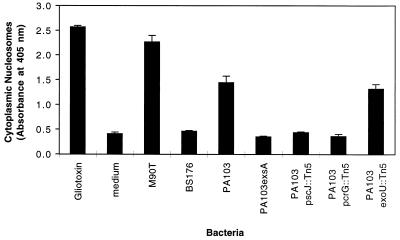
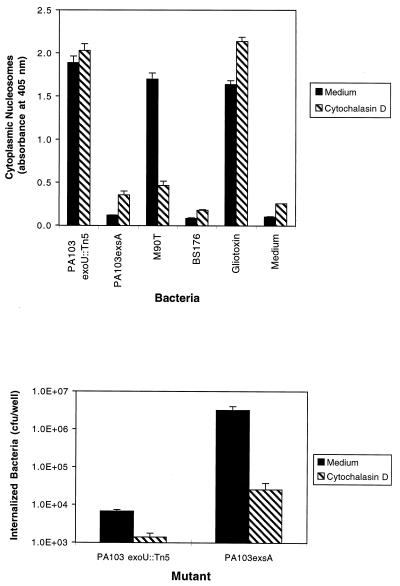
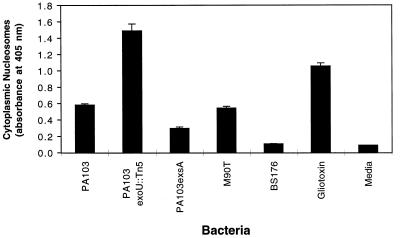
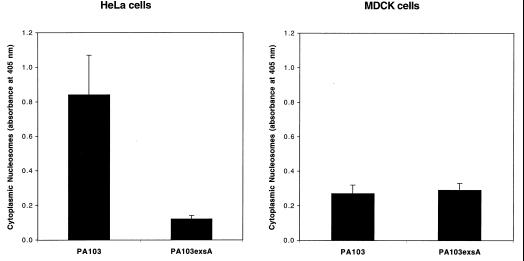
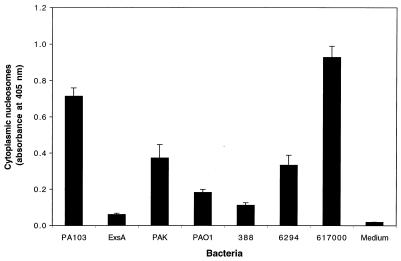
Pseudomonas aeruginosa is a gram-negative opportunistic pathogen that is cytotoxic towards a variety of eukaryotic cells. To investigate the effect of this bacterium on macrophages, we infected J774A.1 cells and primary bone-marrow-derived murine macrophages with the P. aeruginosa strain PA103 in vitro. PA103 caused type-III-secretion-dependent killing of macrophages within 2 h of infection. Only a portion of the killing required the putative cytotoxin ExoU. By three criteria, terminal deoxynucleotidyltransferase-mediated dUTP-biotin nick end labeling assays, cytoplasmic nucleosome assays, and Hoechst staining, the ExoU-independent but type-III-secretion-dependent killing exhibited features of apoptosis. Extracellular bacteria were capable of inducing apoptosis, and some laboratory and clinical isolates of P. aeruginosa induced significantly higher levels of this form of cell death than others. Interestingly, HeLa cells but not Madin-Darby canine kidney cells were susceptible to type-III-secretion-mediated apoptosis under the conditions of these assays. These findings are consistent with a model in which the P. aeruginosa type III secretion system transports at least two factors that kill macrophages: ExoU, which causes necrosis, and a second, as yet unidentified, effector protein, which induces apoptosis. Such killing may contribute to the ability of this organism to persist and disseminate within infected patients.
Figures
Similar articles
-
Pseudomonas aeruginosa cystic fibrosis isolates induce rapid, type III secretion-dependent, but ExoU-independent, oncosis of macrophages and polymorphonuclear neutrophils.Infect Immun. 2000 May;68(5):2916-24. doi: 10.1128/IAI.68.5.2916-2924.2000. Infect Immun. 2000. PMID: 10768989 Free PMC article.
-
ExoU expression by Pseudomonas aeruginosa correlates with acute cytotoxicity and epithelial injury.Mol Microbiol. 1997 Aug;25(3):547-57. doi: 10.1046/j.1365-2958.1997.4891851.x. Mol Microbiol. 1997. PMID: 9302017
-
Apoptosis are induced in J774 macrophages upon phagocytosis and killing of Pseudomonas aeruginosa.Cell Immunol. 2013 Nov-Dec;286(1-2):11-5. doi: 10.1016/j.cellimm.2013.10.006. Epub 2013 Nov 6. Cell Immunol. 2013. PMID: 24270217
-
The role of host cell death in Salmonella infections.Curr Top Microbiol Immunol. 2005;289:131-50. doi: 10.1007/3-540-27320-4_6. Curr Top Microbiol Immunol. 2005. PMID: 15791954 Review.
-
ExoU is a potent intracellular phospholipase.Mol Microbiol. 2004 Sep;53(5):1279-90. doi: 10.1111/j.1365-2958.2004.04194.x. Mol Microbiol. 2004. PMID: 15387809 Review.
Cited by
-
Apoptotic response of Chang cells to infection with Pseudomonas aeruginosa strains PAK and PAO-I: molecular ordering of the apoptosis signaling cascade and role of type IV pili.Infect Immun. 2003 May;71(5):2665-73. doi: 10.1128/IAI.71.5.2665-2673.2003. Infect Immun. 2003. PMID: 12704141 Free PMC article.
-
Characterization of Pseudomonas aeruginosa exoenzyme S as a bifunctional enzyme in J774A.1 macrophages.Infect Immun. 2003 Sep;71(9):5296-305. doi: 10.1128/IAI.71.9.5296-5305.2003. Infect Immun. 2003. PMID: 12933877 Free PMC article.
-
Severe bacteremia results in a loss of hepatic bacterial clearance.Am J Respir Crit Care Med. 2006 Mar 15;173(6):644-52. doi: 10.1164/rccm.200509-1470OC. Epub 2006 Jan 6. Am J Respir Crit Care Med. 2006. PMID: 16399991 Free PMC article.
-
Chronic alcohol exposure renders epithelial cells vulnerable to bacterial infection.PLoS One. 2013;8(1):e54646. doi: 10.1371/journal.pone.0054646. Epub 2013 Jan 24. PLoS One. 2013. PMID: 23358457 Free PMC article.
-
Pathogenicity islands in bacterial pathogenesis.Clin Microbiol Rev. 2004 Jan;17(1):14-56. doi: 10.1128/CMR.17.1.14-56.2004. Clin Microbiol Rev. 2004. PMID: 14726454 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
