

Functions of two types of NADH oxidases in energy metabolism and oxidative stress of Streptococcus mutans
- PMID: 10498705
- PMCID: PMC103620
- DOI: 10.1128/JB.181.19.5940-5947.1999
Functions of two types of NADH oxidases in energy metabolism and oxidative stress of Streptococcus mutans
Abstract
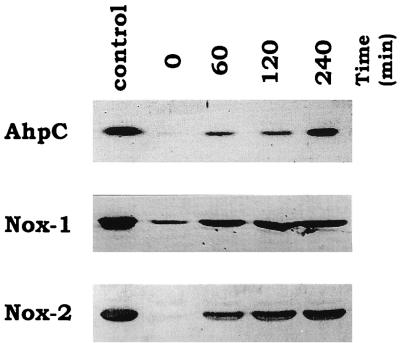
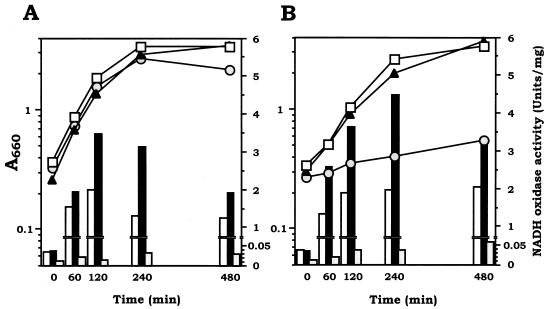
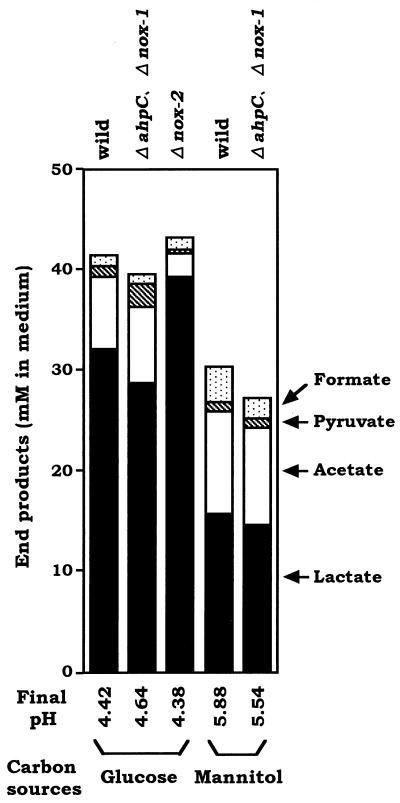
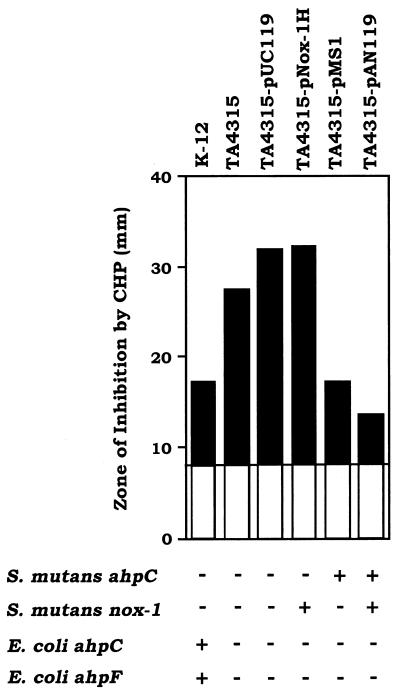
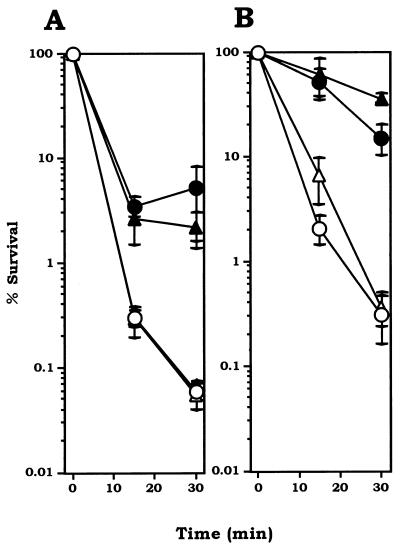
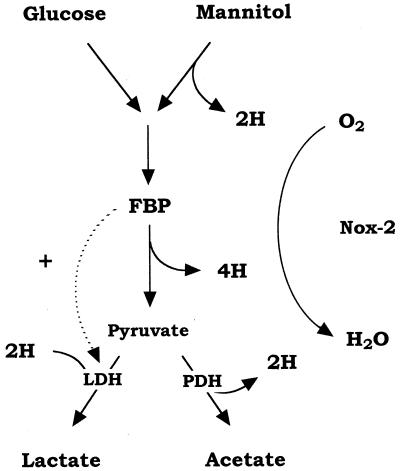
We have previously identified two distinct NADH oxidases corresponding to H(2)O(2)-forming oxidase (Nox-1) and H(2)O-forming oxidase (Nox-2) induced in Streptococcus mutans. Sequence analyses indicated a strong similarity between Nox-1 and AhpF, the flavoprotein component of Salmonella typhimurium alkyl hydroperoxide reductase; an open reading frame upstream of nox-1 also showed homology to AhpC, the direct peroxide-reducing component of S. typhimurium alkyl hydroperoxide reductase. To determine their physiological functions in S. mutans, we constructed knockout mutants of Nox-1, Nox-2, and/or the AhpC homologue; we verified that Nox-2 plays an important role in energy metabolism through the regeneration of NAD(+) but Nox-1 contributes negligibly. The Nox-2 mutant exhibited greatly reduced aerobic growth on mannitol, whereas there was no significant effect of aerobiosis on the growth on mannitol of the other strains or growth on glucose of any of the strains. Although the Nox-2 mutants grew well on glucose aerobically, the end products of glucose fermentation by the Nox-2 mutant were substantially shifted to higher ratios of lactic acid to acetic acid compared with wild-type cells. The resistance to cumene hydroperoxide of Escherichia coli TA4315 (ahpCF-defective mutant) transformed with pAN119 containing both nox-1 and ahpC genes was not only restored but enhanced relative to that of E. coli K-12 (parent strain), indicating a clear function for Nox-1 as part of an alkyl hydroperoxide reductase system in vivo in combination with AhpC. Surprisingly, the Nox-1 and/or AhpC deficiency had no effect on the sensitivity of S. mutans to cumene hydroperoxide and H(2)O(2), implying that the existence of some other antioxidant system(s) independent of Nox-1 in S. mutans compensates for the deficiency.
Figures

References
-
- Bradford M M. A rapid and sensitive methods for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
