

doi: 10.1101/gad.13.18.2360.
Activation of dormant origins of DNA replication in budding yeast
Affiliations
- PMID: 10500092
- PMCID: PMC317032
- DOI: 10.1101/gad.13.18.2360
Item in Clipboard
Activation of dormant origins of DNA replication in budding yeast
Genes Dev.
.
Abstract
Eukaryotic genomes often contain more potential replication origins than are actually used during S phase. The molecular mechanisms that prevent some origins from firing are unknown. Here we show that dormant replication origins on the left arm of budding yeast chromosome III become activated when both passive replication through them is prevented and the Mec1/Rad53 checkpoint that blocks late-origin firing is inactivated. Under these conditions, dormant origins fire very late relative to other active origins. These experiments show that some dormant replication origins are competent to fire during S phase and that passage of a replication fork through such origins can inactivate them.
Figures
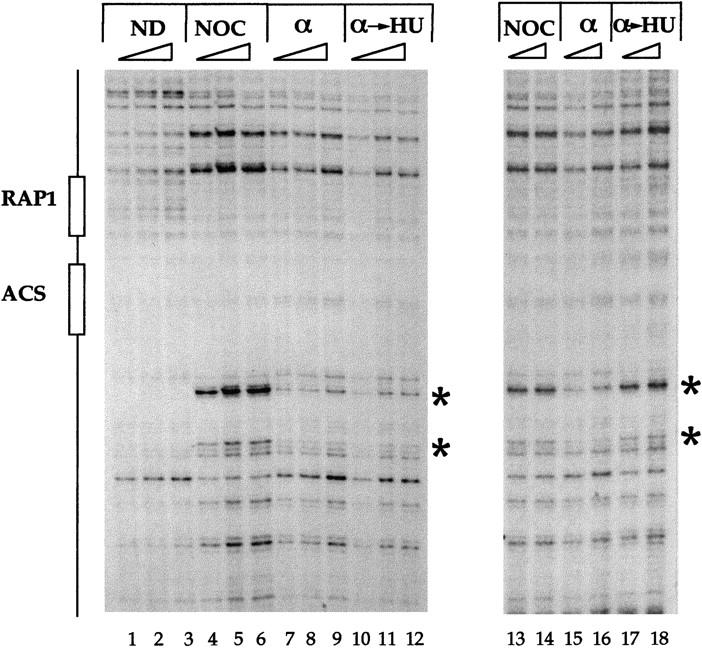
Chromosomal ARS301 conversion from pre- to postreplicative state in HU is blocked by the MEC1/RAD53 checkpoint. Genomic-footprinting analysis of the chromatin structure of ARS301. Wild-type (y300; left) or rad53 (y301; right (Allen et al. 1994) cells were arrested in G2 with nocodazole (NOC) or in G1 with α factor (α) or released from α-factor block into HU for 90 min (α → HU). Naked DNA (ND) was included as control. (*) ORC-induced hypersensitive sites. Cell cycle blocks and genomic footprinting protocols were as described previously (Santocanale and Diffley 1996, 1998). Samples were treated with increasing amounts of DNase I.
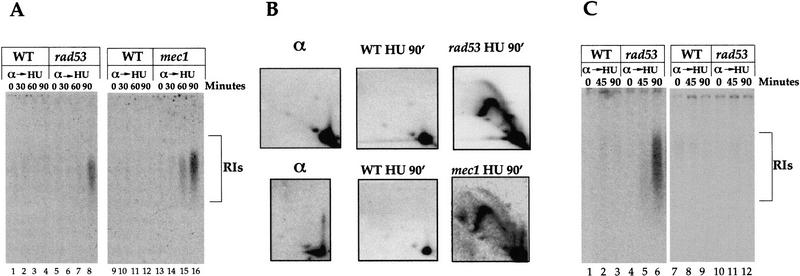
Activation of the dormant chromosomal ARS301 and ARS302/303. (A) Detection of RIs from ARS301. α factor-arrested cells were released from the G1 block into medium containing HU. At the indicated times replication intermediates that originated from ARS301 were studied using the alkaline gel electrophoresis method as described (Santocanale and Diffley 1998). The relevant genotype is indicated. A 207-bp HindIII–BamHI DNA fragment containing the ARS301 sequence prepared from plasmid pCS1 (Santocanale and Diffley 1996) was used as a probe. (B) Characterization of RIs from ARS301. DNAs prepared from α factor-arrested cells (α) or cells released into HU containing medium for 90 min (HU 90′) were analyzed as described in Materials and Methods. (C) Detection of replication intermediates from ARS302. Cells were grown and RIs analyzed as in A. A 1-kb DNA fragment of chromosome III (14,000–15,000 nucleotides) containing the ARS302 sequence was used as a probe for hybridization (lanes 1–6). In lanes 7–12 a 0.8-kb fragment of chromosome III (21,200–22,000 nucleotides) that does not contain origin sequences and lies between ARS303 and ARS304 was used as a probe.
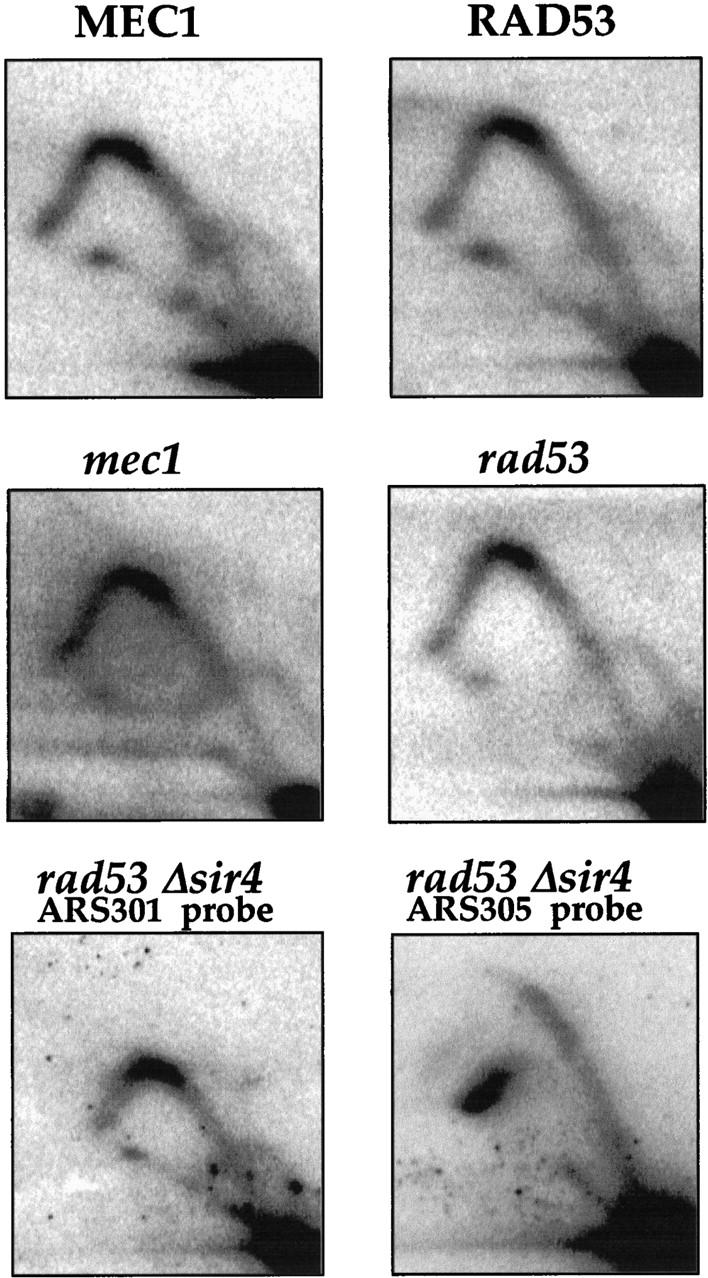
ARS301 is not activated in an unperturbed S phase in checkpoint mutants. Genomic DNA was prepared from logarithmically growing wild-type, rad53, mec1, or rad53/sir4 mutant cells. RIs around ARS301 (or, where indicated, ARS305) were studied as described in Materials and Methods. The relevant genotype of yeast strains Y10860, Y10848 (Paulovich et al. 1997), Y300, Y301 (Allen et al. 1994), and YCS38 (MATa, can1-100, ade2-1, his3-11,15, leu2-3,112, trp1-1, ura3-1, Δsir4::kanR ΔARS1::HIS3, sad1-1) is indicated.
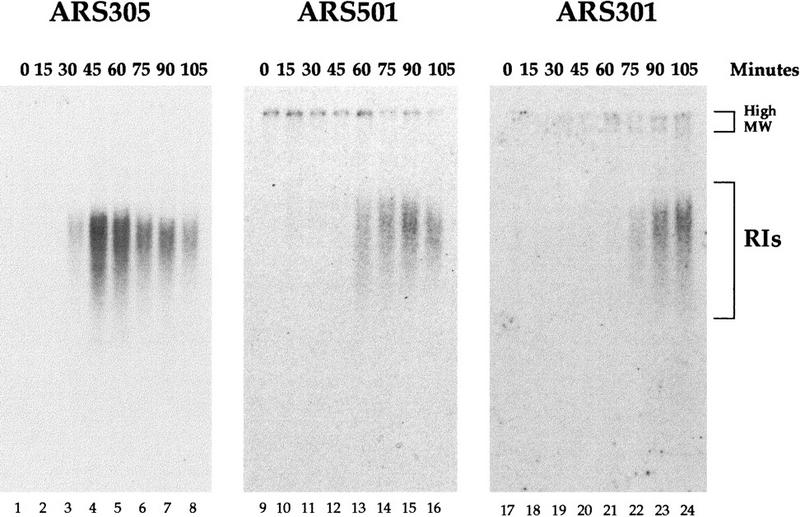
Activation timing of ARS301 in a rad53 mutant in HU. α factor-arrested rad53 cells were released from the G1 block into medium containing HU. At the indicated times aliquots of the culture were collected and genomic DNA prepared as in Fig. 1. Kinetics of RI accumulation from early-firing ARS305, late-firing ARS501, and chromosomal ARS301 were compared using the alkaline gel electrophoresis method.
References
-
- Allen JB, Zhou Z, Siede W, Friedberg EC, Elledge SJ. The SAD1/RAD53 protein kinase controls multiple checkpoints and DNA damage-induced transcription in yeast. Genes & Dev. 1994;8:2401–2415. - PubMed
-
- Blow JJ, Laskey RA. A role for the nuclear envelope in controlling DNA replication within the cell cycle. Nature. 1988;332:546–548. - PubMed
-
- Blumenthal AB, Kriegstein HJ, Hogness DS. The units of DNA replication in Drosophila melanogaster chromosomes. Cold Spring Harb Symp Quant Biol. 1973;38:205–223. - PubMed
-
- Brewer BJ, Fangman WL. The localization of replication origins on ARS plasmids in S. cerevisiae. Cell. 1987;51:463–471. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases