

doi: 10.1101/gad.13.18.2449.
The Escherichia coli sigma(E)-dependent extracytoplasmic stress response is controlled by the regulated proteolysis of an anti-sigma factor
Affiliations
- PMID: 10500101
- PMCID: PMC317020
- DOI: 10.1101/gad.13.18.2449
Item in Clipboard
The Escherichia coli sigma(E)-dependent extracytoplasmic stress response is controlled by the regulated proteolysis of an anti-sigma factor
Genes Dev.
.
Abstract
The activity of the stress-responsive sigma factor, sigma(E), is induced by the extracytoplasmic accumulation of misfolded or unfolded protein. The inner membrane protein RseA is the central regulatory molecule in this signal transduction cascade and acts as a sigma(E)-specific anti-sigma factor. Here we show that sigma(E) activity is primarily determined by the ratio of RseA to sigma(E). RseA is rapidly degraded in response to extracytoplasmic stress, leading to an increase in the free pool of sigma(E) and initiation of the stress response. We present evidence that the putative inner membrane serine protease, DegS, is responsible for this regulated degradation of RseA.
Figures
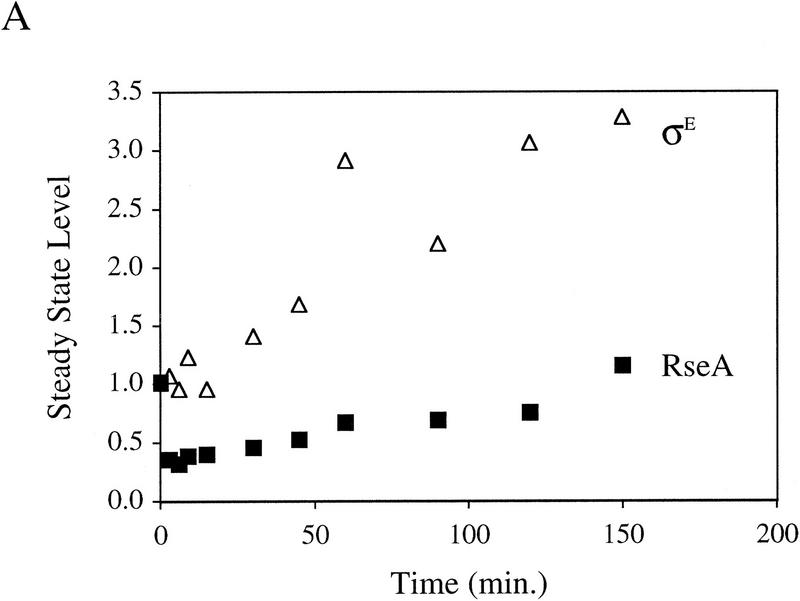
Steady-state levels of σE and RseA, and EσE activity during the stress response. (A) Steady-state levels of RseA and σE during the extracytoplasmic stress response. Wild-type cells containing pompC (CAG45025) were grown to early-log phase and IPTG added at t = 0 to induce overexpression of OmpC. The levels of RseA (█) and σE (▵) were determined by Western blot analysis as described in Materials and Methods. The amount of each protein at a given time was normalized to the amount of that protein at time t = 0. A representative data set is shown. (B) EσE activity during the extracytoplasmic stress response. Wild-type cells containing pompC (CAG45025) were grown to early-log phase and IPTG was added at t = 0. EσE activity was determined at various time points by measuring the rate of synthesis of β-galactosidase (○) from a single-copy σE-dependent lacZ reporter gene in Φλ[rpoHP3–lacZ] using the pulse-labeling protocol described in Materials and Methods. The synthesis rate shown for β-galactosidase is normalized to the synthesis rate of the protein at t = 0. A representative data set is shown.
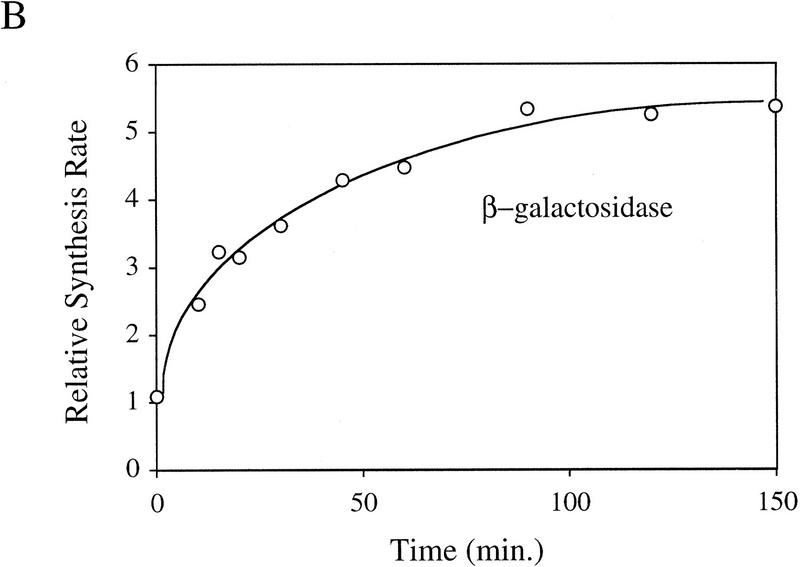
Steady-state levels of σE and RseA, and EσE activity during the stress response. (A) Steady-state levels of RseA and σE during the extracytoplasmic stress response. Wild-type cells containing pompC (CAG45025) were grown to early-log phase and IPTG added at t = 0 to induce overexpression of OmpC. The levels of RseA (█) and σE (▵) were determined by Western blot analysis as described in Materials and Methods. The amount of each protein at a given time was normalized to the amount of that protein at time t = 0. A representative data set is shown. (B) EσE activity during the extracytoplasmic stress response. Wild-type cells containing pompC (CAG45025) were grown to early-log phase and IPTG was added at t = 0. EσE activity was determined at various time points by measuring the rate of synthesis of β-galactosidase (○) from a single-copy σE-dependent lacZ reporter gene in Φλ[rpoHP3–lacZ] using the pulse-labeling protocol described in Materials and Methods. The synthesis rate shown for β-galactosidase is normalized to the synthesis rate of the protein at t = 0. A representative data set is shown.
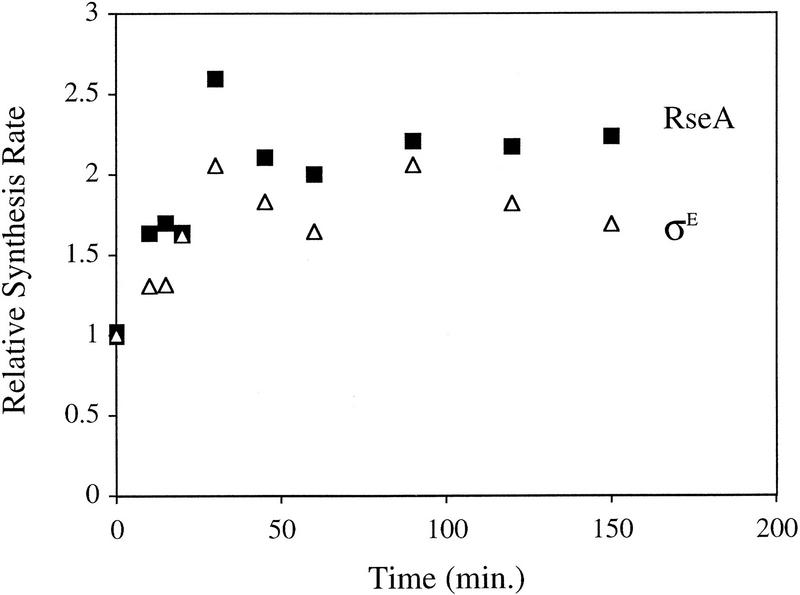
Synthesis of σE and RseA during the extracytoplasmic stress response, induced in wild-type cells containing pompC (CAG45025) by the addition of IPTG at t = 0. The rates of synthesis of σE (▵) and RseA (█) at various time points after induction were determined by pulse labeling as described in Materials and Methods. The rate at each time point for a given protein was normalized to the initial rate for that protein at t = 0. A representative data set is shown.
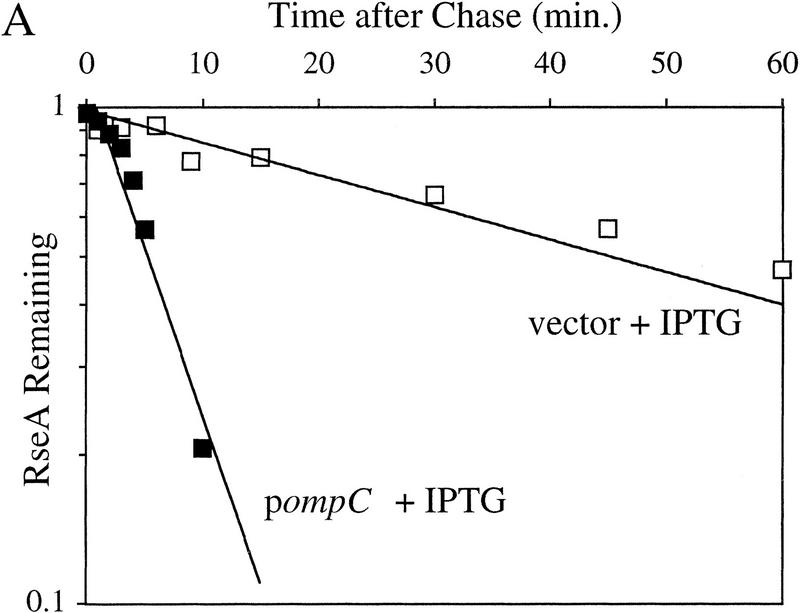
RseA stability under stress and nonstress conditions. (A) The stability of RseA synthesized after initiation of the stress response in cells overexpressing OmpC. Wild-type cells containing pompC (CAG45025, █) or vector alone (CAG45027, □) were grown to early log phase, and the stress response was initiated by the addition of IPTG to induce expression of OmpC. After 10 min of induction, the cells were pulse-labeled with [35S]methionine followed by a chase of cold methionine and cysteine. The stability of RseA was determined as described in Materials and Methods. A representative data set is shown. (B) The stability of RseA synthesized after temperature upshift. Wild-type cells (CAG16037) were grown to early log phase and either left at 30°C (⋄) or shifted to 43°C (♦). Five minutes after shift to high temperature the cells were pulse-labeled with [35S]methionine, followed by a chase of cold methionine and cysteine. The stability of RseA was determined as described in Materials and Methods. A representative data set is shown.
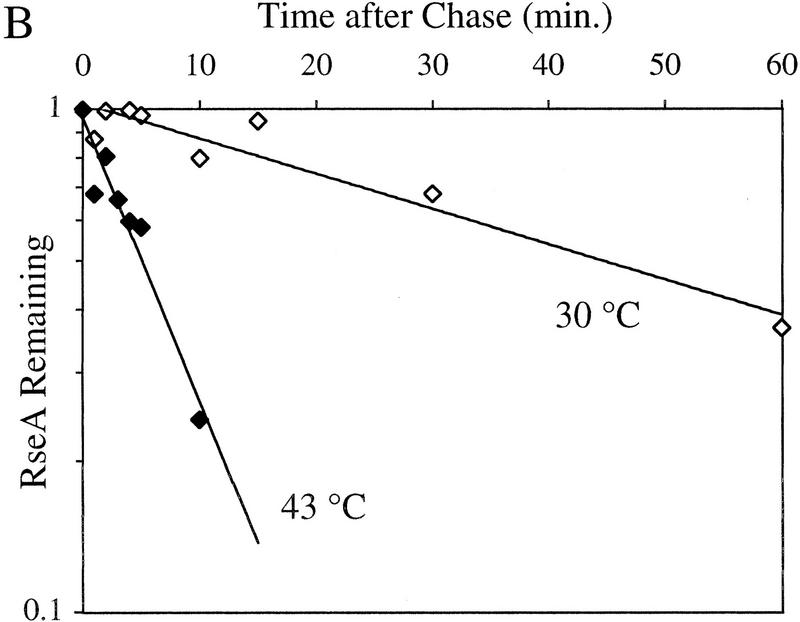
RseA stability under stress and nonstress conditions. (A) The stability of RseA synthesized after initiation of the stress response in cells overexpressing OmpC. Wild-type cells containing pompC (CAG45025, █) or vector alone (CAG45027, □) were grown to early log phase, and the stress response was initiated by the addition of IPTG to induce expression of OmpC. After 10 min of induction, the cells were pulse-labeled with [35S]methionine followed by a chase of cold methionine and cysteine. The stability of RseA was determined as described in Materials and Methods. A representative data set is shown. (B) The stability of RseA synthesized after temperature upshift. Wild-type cells (CAG16037) were grown to early log phase and either left at 30°C (⋄) or shifted to 43°C (♦). Five minutes after shift to high temperature the cells were pulse-labeled with [35S]methionine, followed by a chase of cold methionine and cysteine. The stability of RseA was determined as described in Materials and Methods. A representative data set is shown.
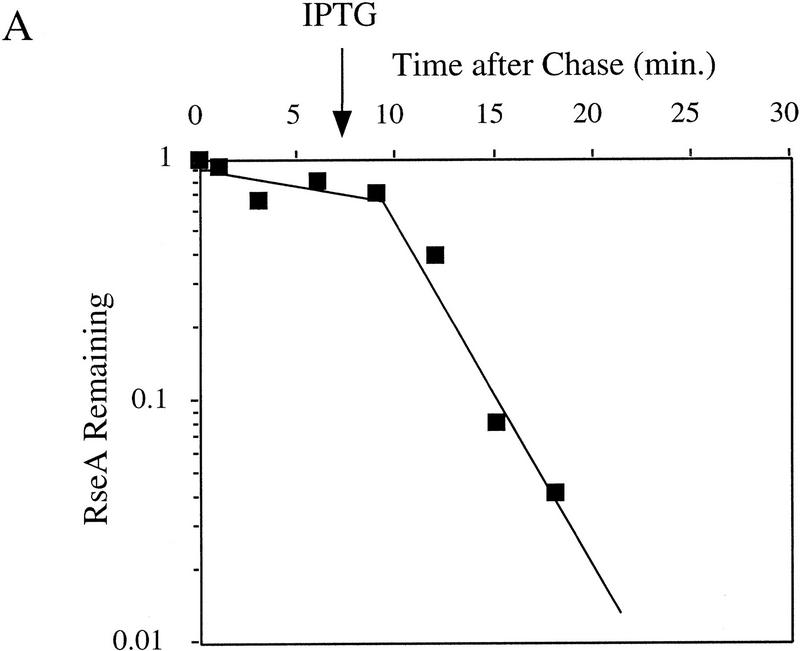
The stability of RseA synthesized prior to initiation of the stress response. Cells containing pompC (CAG45025, █) or wild-type cells (CAG16037, ♦) were grown to early log phase and pulse-labeled with [35S]methionine, followed by a chase of cold methionine and cysteine. At the indicated times, IPTG was added to induce overexpression of OmpC (A) or the cells were shifted from 30°C to 43°C (B). The stability of RseA was determined as described in Materials and Methods. A representative data set is shown.
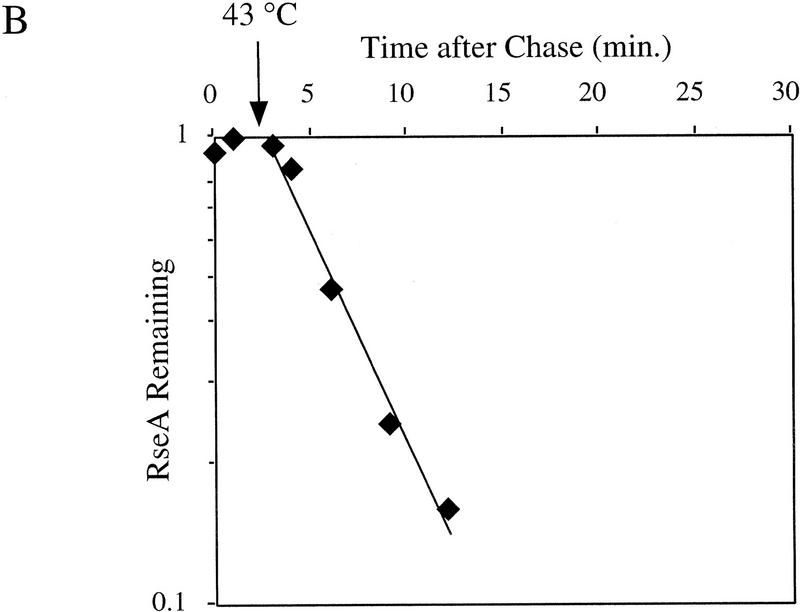
The stability of RseA synthesized prior to initiation of the stress response. Cells containing pompC (CAG45025, █) or wild-type cells (CAG16037, ♦) were grown to early log phase and pulse-labeled with [35S]methionine, followed by a chase of cold methionine and cysteine. At the indicated times, IPTG was added to induce overexpression of OmpC (A) or the cells were shifted from 30°C to 43°C (B). The stability of RseA was determined as described in Materials and Methods. A representative data set is shown.
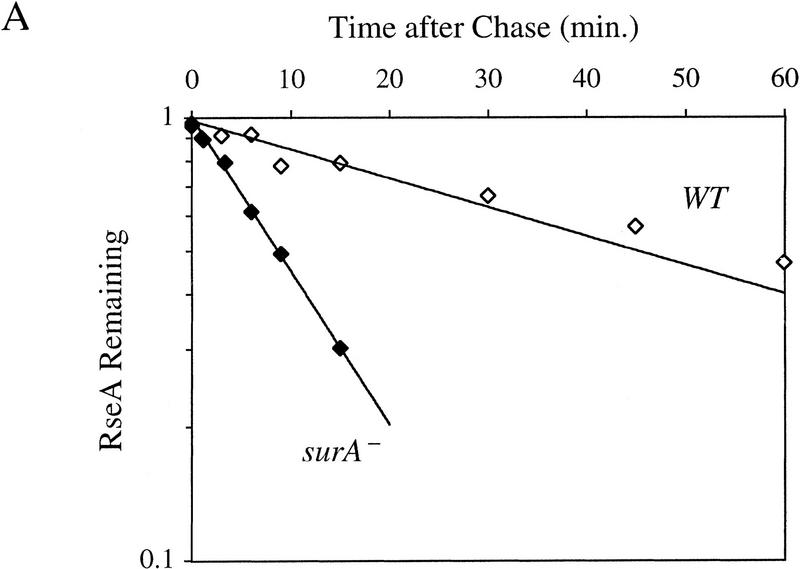
RseA stability under conditions of constitutive activation of σE. (A) RseA stability in cells lacking the periplasmic peptidyl prolyl isomerase SurA. surA− (CAG24029, ♦) or isogenic wild-type cells (CAG16037, ⋄) were grown to early log phase and then pulse-labeled with [35S]methionine, followed by a chase of cold methionine and cysteine. The stability of RseA was then determined as described in Materials and Methods. A representative data set is shown. (B) RseA stability in cells lacking RseB. ΔrseB cells (CAG22951, ♦) or isogenic wild-type cells (CAG16037, ⋄) were grown to early log phase and then pulse-labeled with [35S]methionine, followed by a chase of cold methionine and cysteine. The stability of RseA was then determined as described in Materials and Methods. A representative data set is shown.
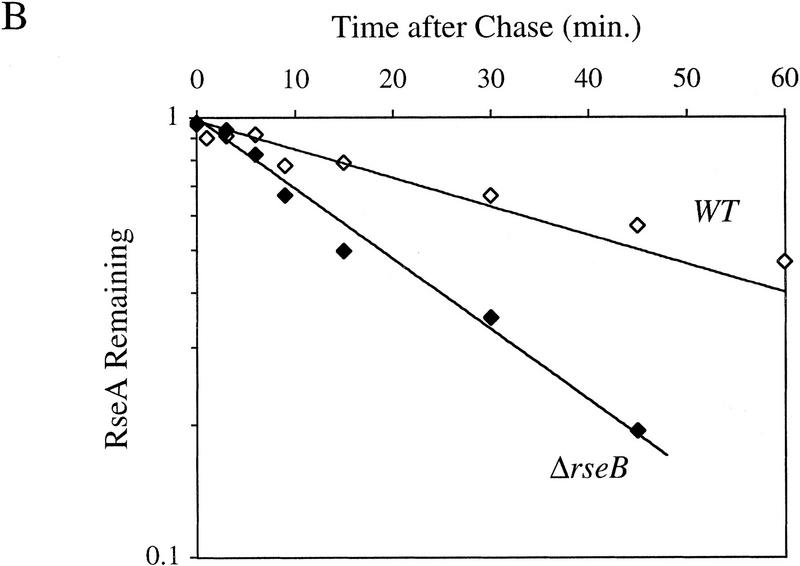
RseA stability under conditions of constitutive activation of σE. (A) RseA stability in cells lacking the periplasmic peptidyl prolyl isomerase SurA. surA− (CAG24029, ♦) or isogenic wild-type cells (CAG16037, ⋄) were grown to early log phase and then pulse-labeled with [35S]methionine, followed by a chase of cold methionine and cysteine. The stability of RseA was then determined as described in Materials and Methods. A representative data set is shown. (B) RseA stability in cells lacking RseB. ΔrseB cells (CAG22951, ♦) or isogenic wild-type cells (CAG16037, ⋄) were grown to early log phase and then pulse-labeled with [35S]methionine, followed by a chase of cold methionine and cysteine. The stability of RseA was then determined as described in Materials and Methods. A representative data set is shown.
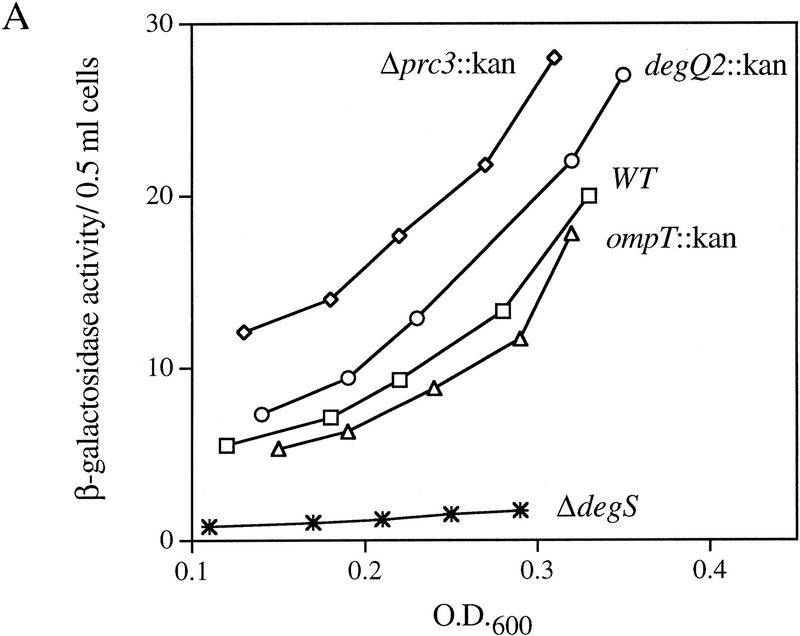
EσE activity in cells lacking various extracytoplasmic proteases. (A) EσE activity in strains lacking the extracytoplasmic proteases Prc (Tsp), DegQ, OmpT, or DegS. EσE activity was assayed by monitoring β-galactosidase activity produced from a single copy Φλ[rpoHP3–lacZ] fusion in wild-type (CAG16037, □), and the isogenic Δprc3::Km (CAG33314, ⋄), degQ2::Km (CAG33397, ○), ompT::Km (CAG33263, ▵), and ΔdegS (CAG33315, ✠ ) strains. β-Galactosidase assays were performed as described in Materials and Methods. A representative data set is shown. (B) EσE activity in cells lacking DegS. EσE activity was assayed in wild-type (CAG33385, □) and isogenic ΔdegS (CAG33391, ⋄) strains carrying vector alone as well as in ΔdegS cells carrying the plasmid pdegS (CAG33393, ○) or pdegS S201A (CAG33382, ▵). A representative data set is shown.
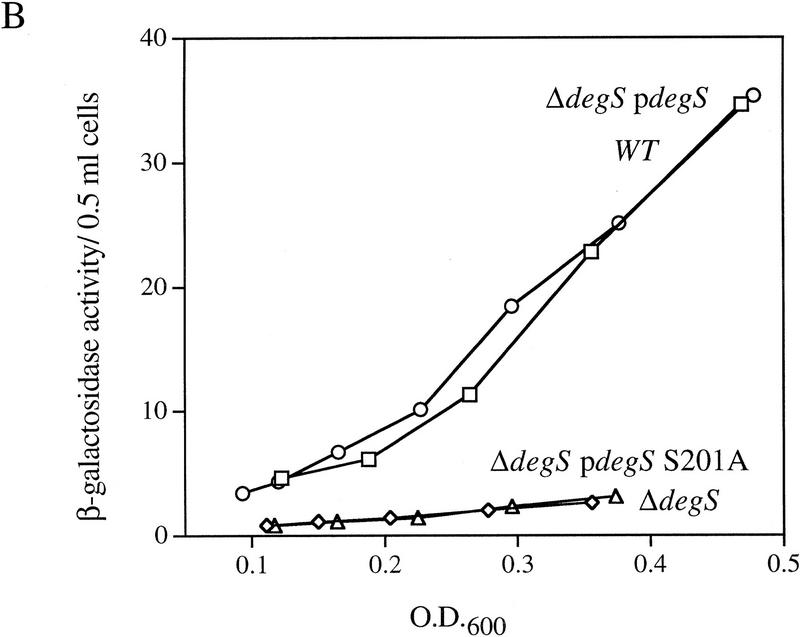
EσE activity in cells lacking various extracytoplasmic proteases. (A) EσE activity in strains lacking the extracytoplasmic proteases Prc (Tsp), DegQ, OmpT, or DegS. EσE activity was assayed by monitoring β-galactosidase activity produced from a single copy Φλ[rpoHP3–lacZ] fusion in wild-type (CAG16037, □), and the isogenic Δprc3::Km (CAG33314, ⋄), degQ2::Km (CAG33397, ○), ompT::Km (CAG33263, ▵), and ΔdegS (CAG33315, ✠ ) strains. β-Galactosidase assays were performed as described in Materials and Methods. A representative data set is shown. (B) EσE activity in cells lacking DegS. EσE activity was assayed in wild-type (CAG33385, □) and isogenic ΔdegS (CAG33391, ⋄) strains carrying vector alone as well as in ΔdegS cells carrying the plasmid pdegS (CAG33393, ○) or pdegS S201A (CAG33382, ▵). A representative data set is shown.
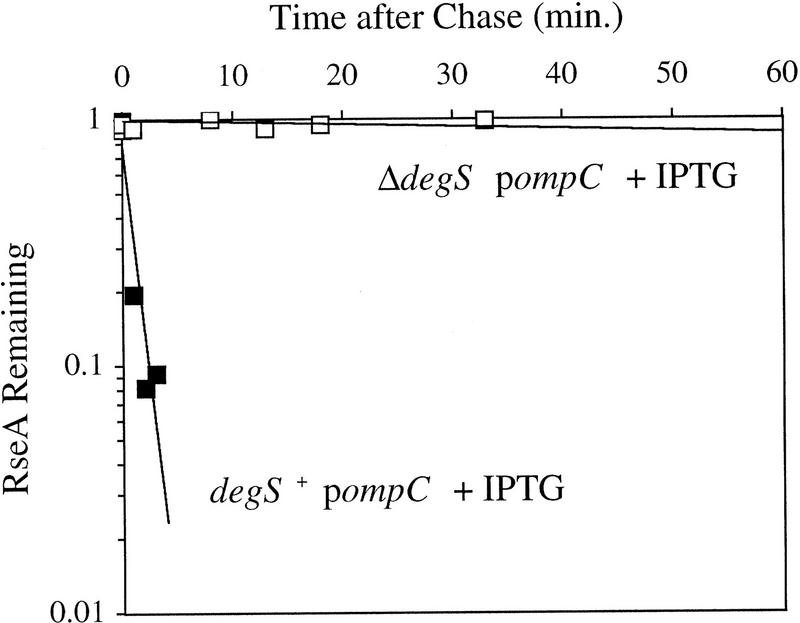
RseA stability in cells lacking DegS. sup degS+ (CAG41073, █) or sup ΔdegS (CAG43150, □) cells were grown to early log phase, and the stress response was initiated by the addition of IPTG to induce expression of OmpC. Five minutes after induction of the stress response the cells were pulse-labeled with [35S]methionine, followed by a chase of cold methionine and cysteine. The stability of RseA was determined as described in Materials and Methods. A representative data set is shown. The stability of RseA in sup-containing cells is greater than that in wild-type cells, consistent with the lower basal activity of σE in sup-containing cells (data not shown).
References
-
- Akiyama Y, Ito K. SecY protein, a membrane embedded secretion factor of E. coli, is cleaved by the OmpT protease in vitro. Biochem Biophys Res Commun. 1996;167:711–715. - PubMed
-
- Bartolomé B, Jubete Y, Martinez E, de la Cruz F. Construction and properties of a family of pACYC184-derived cloning vectors compatible with pBR322 and its derivatives. Gene. 1991;102:75–78. - PubMed
-
- Becker J, Craig EA. Heat-shock proteins as molecular chaperones. Eur J Biochem. 1994;219:11–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases