

Circular permutation and receptor insertion within green fluorescent proteins
- PMID: 10500161
- PMCID: PMC18018
- DOI: 10.1073/pnas.96.20.11241
Circular permutation and receptor insertion within green fluorescent proteins
Abstract
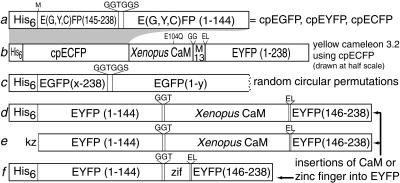
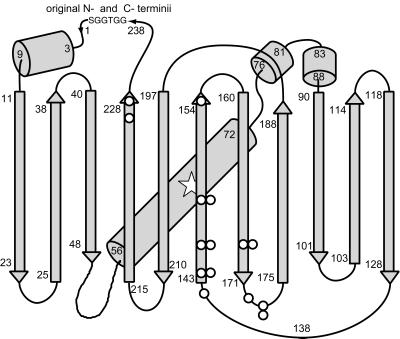
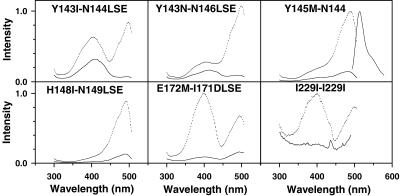
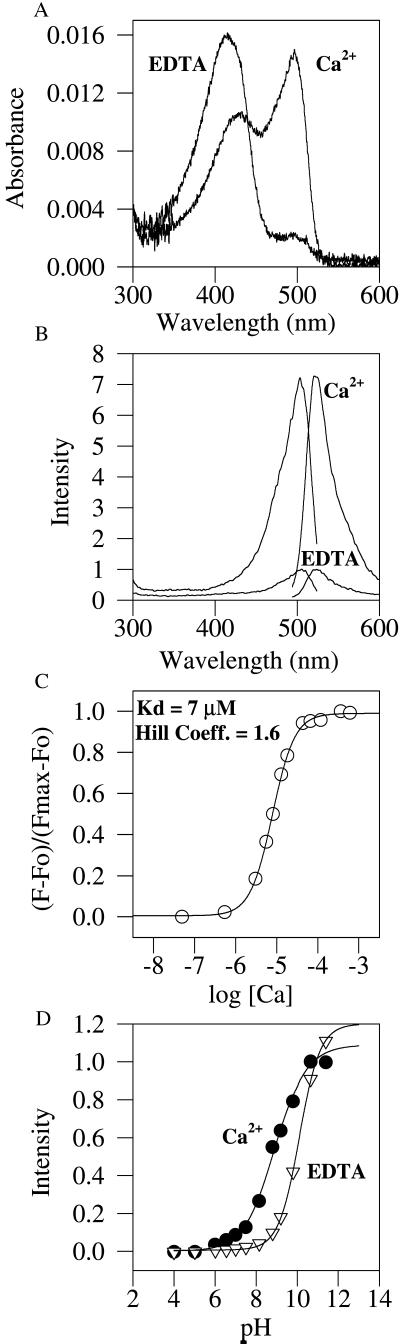
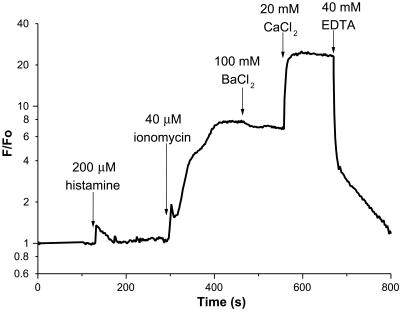
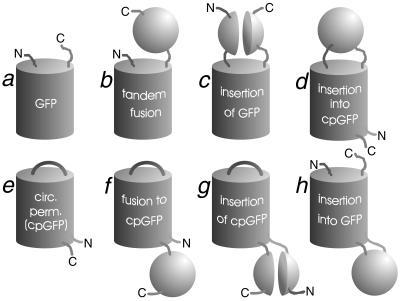
Many areas of biology and biotechnology have been revolutionized by the ability to label proteins genetically by fusion to the Aequorea green fluorescent protein (GFP). In previous fusions, the GFP has been treated as an indivisible entity, usually appended to the amino or carboxyl terminus of the host protein, occasionally inserted within the host sequence. The tightly interwoven, three-dimensional structure and intricate posttranslational self-modification required for chromophore formation would suggest that major rearrangements or insertions within GFP would prevent fluorescence. However, we now show that several rearrangements of GFPs, in which the amino and carboxyl portions are interchanged and rejoined with a short spacer connecting the original termini, still become fluorescent. These circular permutations have altered pKa values and orientations of the chromophore with respect to a fusion partner. Furthermore, certain locations within GFP tolerate insertion of entire proteins, and conformational changes in the insert can have profound effects on the fluorescence. For example, insertions of calmodulin or a zinc finger domain in place of Tyr-145 of a yellow mutant (enhanced yellow fluorescent protein) of GFP result in indicator proteins whose fluorescence can be enhanced severalfold upon metal binding. The calmodulin graft into enhanced yellow fluorescent protein can monitor cytosolic Ca(2+) in single mammalian cells. The tolerance of GFPs for circular permutations and insertions shows the folding process is surprisingly robust and offers a new strategy for creating genetically encodable, physiological indicators.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
