

Sulfation of a high endothelial venule-expressed ligand for L-selectin. Effects on tethering and rolling of lymphocytes
- PMID: 10510083
- PMCID: PMC2195654
- DOI: 10.1084/jem.190.7.935
Sulfation of a high endothelial venule-expressed ligand for L-selectin. Effects on tethering and rolling of lymphocytes
Abstract
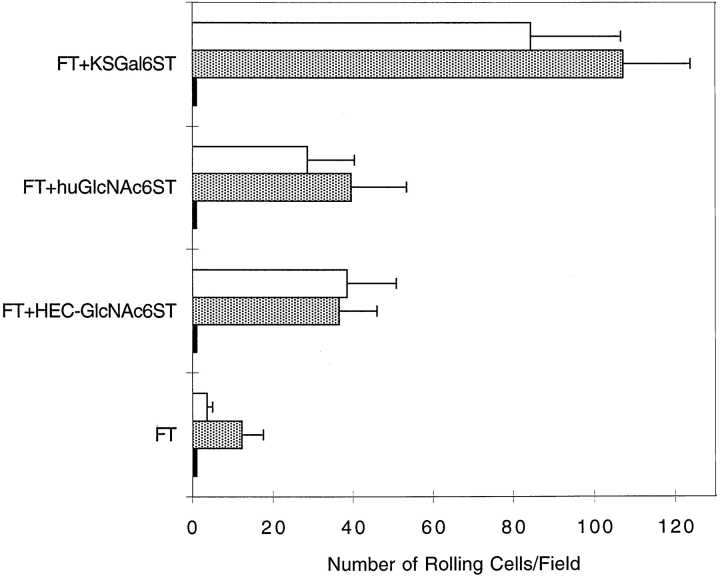
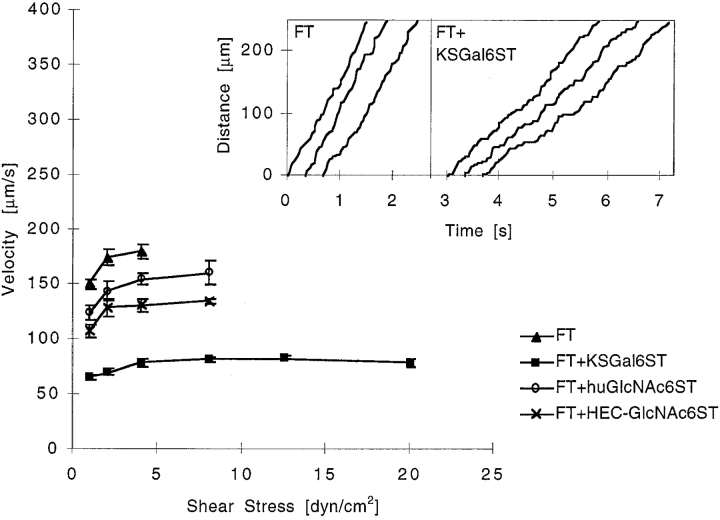
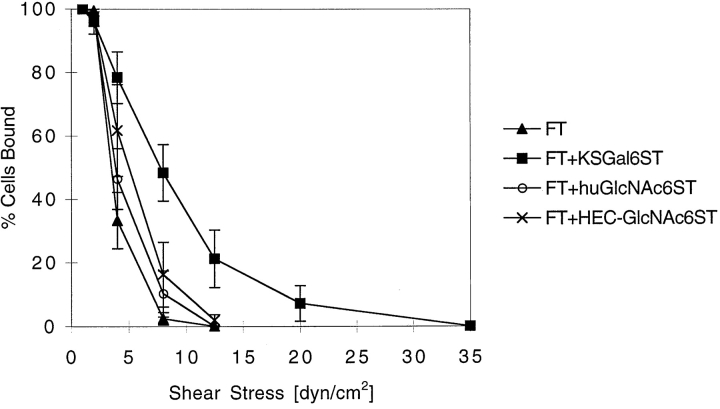
During lymphocyte homing, L-selectin mediates the tethering and rolling of lymphocytes on high endothelial venules (HEVs) in secondary lymphoid organs. The L-selectin ligands on HEV are a set of mucin-like glycoproteins, for which glycosylation-dependent cell adhesion molecule 1 (GlyCAM-1) is a candidate. Optimal binding in equilibrium measurements requires sulfation, sialylation, and fucosylation of ligands. Analysis of GlyCAM-1 has revealed two sulfation modifications (galactose [Gal]-6-sulfate and N-acetylglucosamine [GlcNAc]-6-sulfate) of sialyl Lewis x. Recently, three related sulfotransferases (keratan sulfate galactose-6-sulfotransferase [KSGal6ST], high endothelial cell N-acetylglucosamine-6-sulfotransferase [GlcNAc6ST], and human GlcNAc6ST) were cloned, which can generate Gal-6-sulfate and GlcNAc-6-sulfate in GlyCAM-1. Imparting these modifications to GlyCAM-1, together with appropriate fucosylation, yields enhanced rolling ligands for both peripheral blood lymphocytes and Jurkat cells in flow chamber assays as compared with those generated with exogenous fucosyltransferase. Either sulfation modification results in an increased number of tethered and rolling lymphocytes, a reduction in overall rolling velocity associated with more frequent pausing of the cells, and an enhanced resistance of rolling cells to detachment by shear. All of these effects are predicted to promote the overall efficiency of lymphocyte homing. In contrast, the rolling interactions of E-selectin transfectants with the same ligands are not affected by sulfation.
Figures



References
-
- Girard J.-P., Springer T.A. High endothelial venulesspecialized endothelium for lymphocyte migration. Immunol. Today. 1995;16:449–457. - PubMed
-
- Butcher E.C., Picker L.J. Lymphocyte homing and homeostasis. Science. 1996;272:60–66. - PubMed
-
- Rosen S.D. Selectins. In: Vale R., Kreis T., editors. Guide to the Extracellular Matrix and Adhesion Proteins. 2nd Edition. Oxford University Press; Oxford, UK: 1999. pp. 290–297.
-
- Imai Y., Lasky L.A., Rosen S.D. Sulphation requirement for GlyCAM-1, an endothelial ligand for L-selectin. Nature. 1993;361:555–557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
