

Defective development of gamma/delta T cells in interleukin 7 receptor-deficient mice is due to impaired expression of T cell receptor gamma genes
- PMID: 10510087
- PMCID: PMC2195640
- DOI: 10.1084/jem.190.7.973
Defective development of gamma/delta T cells in interleukin 7 receptor-deficient mice is due to impaired expression of T cell receptor gamma genes
Abstract
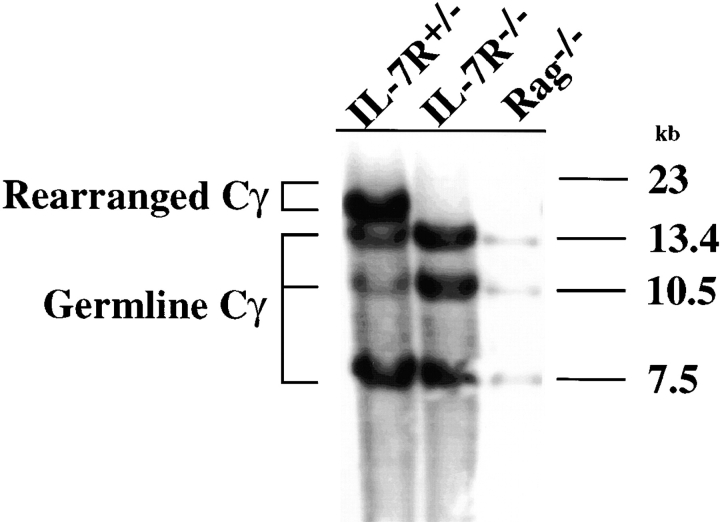
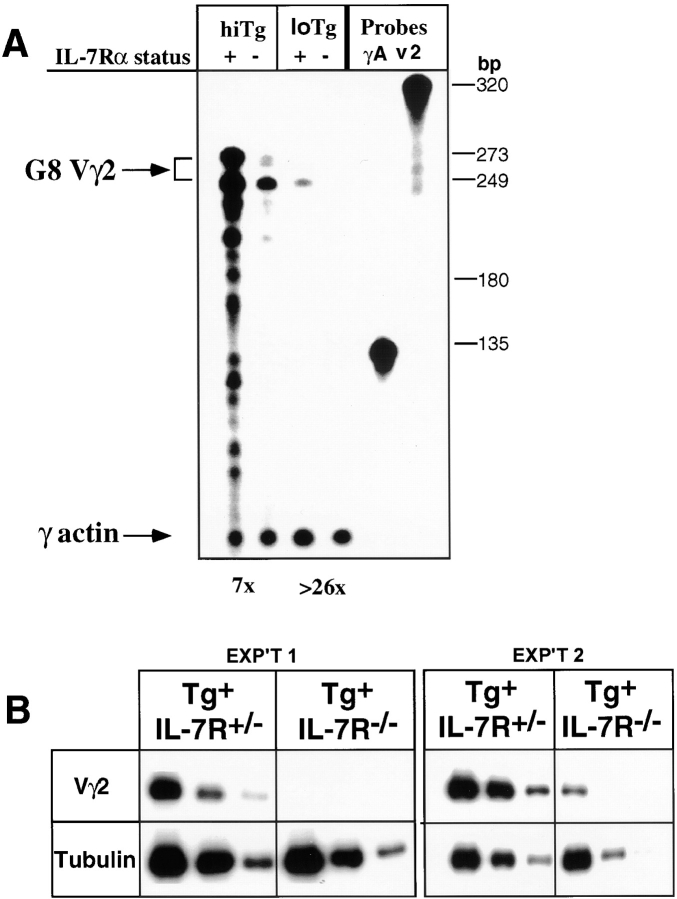
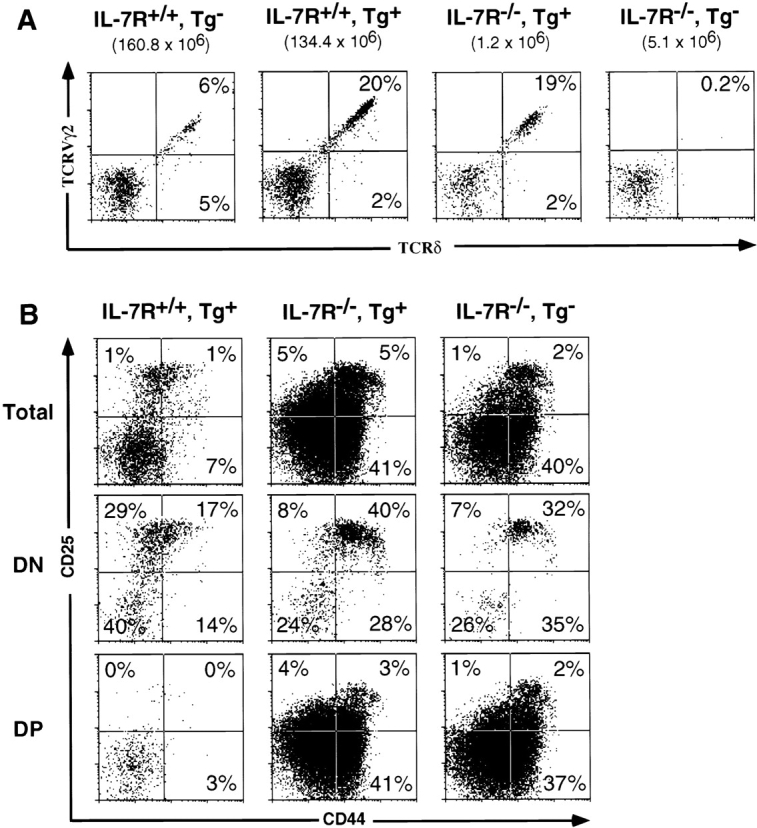
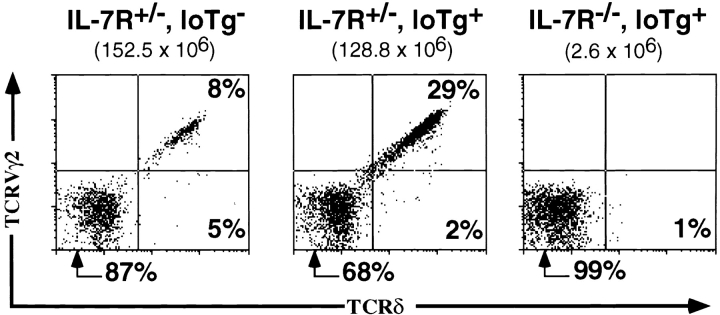
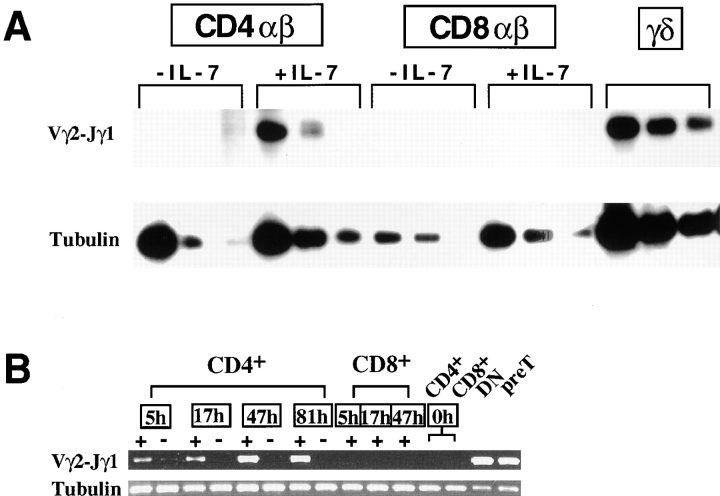
Mice lacking the interleukin 7 receptor (IL-7R) generate alpha/beta T cells at a detectable but greatly reduced rate, but gamma/delta T cells are completely absent. The special role of IL-7R signaling in gamma/delta T cell development has remained unclear. IL-7Ralpha(-/-) mice exhibit a paucity of gamma gene rearrangements. This striking observation can be explained by a defect in T cell receptor (TCR)-gamma gene rearrangement, a defect in TCR-gamma gene transcription leading to death of gamma/delta lineage cells, and/or a requirement for IL-7R in commitment of cells to the gamma/delta lineage. To determine the role of IL-7R signaling in gamma/delta T cell development, we examined transcription of a prerearranged TCR-gamma transgene in IL-7Ralpha(-/-) mice, as well as the effects of IL-7 on transcription of endogenous, rearranged TCR-gamma genes in alpha/beta lineage cells. The results demonstrate that IL-7R-mediated signals are necessary for the normal expression of rearranged TCR-gamma genes. Equally significant, the results show that the poor expression of TCR-gamma genes in IL-7Ralpha(-/-) mice is responsible for the selective deficit in gamma/delta cells in these mice, since a high copy TCR-gamma transgene exhibited sufficient residual expression in IL-7Ralpha(-/-) mice to drive gamma/delta cell development. The results indicate that the absence of gamma/delta T cells in IL-7Ralpha(-/-) mice is due to insufficient TCR-gamma gene expression.
Figures
Similar articles
-
Interleukin 7 receptor control of T cell receptor gamma gene rearrangement: role of receptor-associated chains and locus accessibility.J Exp Med. 1998 Dec 21;188(12):2233-41. doi: 10.1084/jem.188.12.2233. J Exp Med. 1998. PMID: 9858510 Free PMC article.
-
The interleukin 7 receptor is required for T cell receptor gamma locus accessibility to the V(D)J recombinase.J Exp Med. 2000 Mar 20;191(6):1045-50. doi: 10.1084/jem.191.6.1045. J Exp Med. 2000. PMID: 10727465 Free PMC article.
-
Defective T-cell receptor gamma gene rearrangement in interleukin-7 receptor knockout mice.Immunol Lett. 1997 Jun 1;57(1-3):9-14. doi: 10.1016/s0165-2478(97)00062-x. Immunol Lett. 1997. PMID: 9232418
-
Crucial function of the pre-T-cell receptor (TCR) in TCR beta selection, TCR beta allelic exclusion and alpha beta versus gamma delta lineage commitment.Immunol Rev. 1998 Oct;165:111-9. doi: 10.1111/j.1600-065x.1998.tb01234.x. Immunol Rev. 1998. PMID: 9850856 Review.
-
Role of the IL-7 receptor in gamma-delta T cell development.Chem Immunol. 2001;79:29-42. doi: 10.1159/000058830. Chem Immunol. 2001. PMID: 11478153 Review. No abstract available.
Cited by
-
Evidence that gammadelta versus alphabeta T cell fate determination is initiated independently of T cell receptor signaling.J Exp Med. 2001 Mar 19;193(6):689-98. doi: 10.1084/jem.193.6.689. J Exp Med. 2001. PMID: 11257136 Free PMC article.
-
Fine-scale staging of T cell lineage commitment in adult mouse thymus.J Immunol. 2010 Jul 1;185(1):284-93. doi: 10.4049/jimmunol.1000679. Epub 2010 Jun 11. J Immunol. 2010. PMID: 20543111 Free PMC article.
-
Cytokine signal transduction is suppressed in preselection double-positive thymocytes and restored by positive selection.J Exp Med. 2006 Jan 23;203(1):165-75. doi: 10.1084/jem.20051836. Epub 2006 Jan 3. J Exp Med. 2006. PMID: 16390939 Free PMC article.
-
Harnessing the Power of IL-7 to Boost T Cell Immunity in Experimental and Clinical Immunotherapies.Immune Netw. 2024 Feb 15;24(1):e9. doi: 10.4110/in.2024.24.e9. eCollection 2024 Feb. Immune Netw. 2024. PMID: 38455462 Free PMC article. Review.
-
From thymus to periphery: Molecular basis of effector γδ-T cell differentiation.Immunol Rev. 2020 Nov;298(1):47-60. doi: 10.1111/imr.12918. Epub 2020 Nov 15. Immunol Rev. 2020. PMID: 33191519 Free PMC article. Review.
References
-
- Wiles M.V., Ruiz P., Imhof B.A. Interleukin-7 expression during mouse thymus development. Eur. J. Immunol. 1992;22:1037–1042. - PubMed
-
- Kondo M., Weissman I.L., Akashi K. Identification of clonogenic common lymphoid progenitors in mouse bone marrow. Cell. 1997;91:661–672. - PubMed
-
- Lin J.X., Migone T.S., Tsang M., Friedmann M., Weatherbee J.A., Zhou L., Yamauchi A., Bloom E.T., Mietz J., John S. The role of shared receptor motifs and common Stat proteins in the generation of cytokine pleiotropy and redundancy by IL-2, IL-4, IL-7, IL-13, and IL-15. Immunity. 1995;2:331–339. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
