

Epithelial P2X purinergic receptor channel expression and function
- PMID: 10510328
- PMCID: PMC408558
- DOI: 10.1172/JCI7270
Epithelial P2X purinergic receptor channel expression and function
Abstract
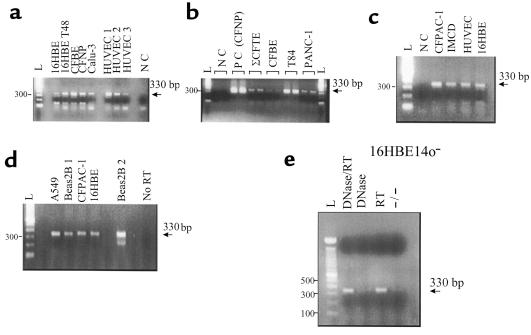
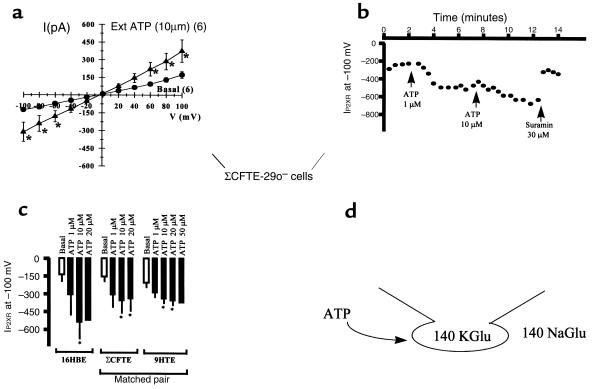
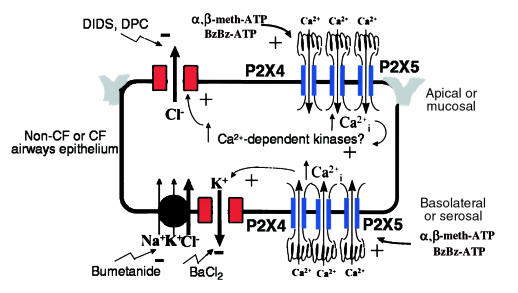
P2X purinergic receptor (P2XR) channels bind ATP and mediate Ca(2+) influx--2 signals that stimulate secretory Cl(-) transport across epithelia. We tested the hypotheses that P2XR channels are expressed by epithelia and that P2XRs transduce extracellular ATP signals into stimulation of Cl(-) transport across epithelia. Electrophysiological data and mRNA analysis of human and mouse pulmonary epithelia and other epithelial cells indicate that multiple P2XRs are broadly expressed in these tissues and that they are active on both apical and basolateral surfaces. Because P2X-selective agonists bind multiple P2XR subtypes, and because P2X agonists stimulate Cl(-) transport across nasal mucosa of cystic fibrosis (CF) patients as well as across non-CF nasal mucosa, P2XRs may provide novel targets for extracellular nucleotide therapy of CF.
Figures
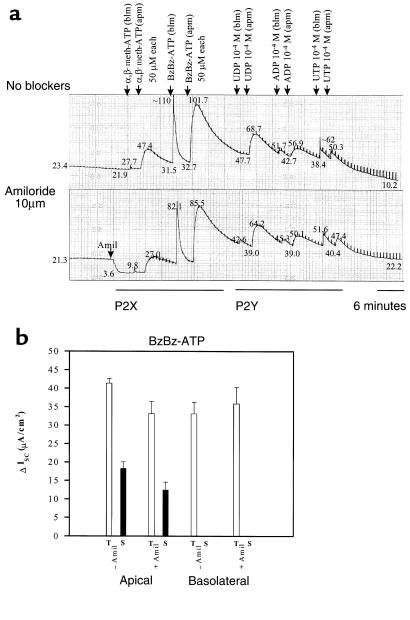
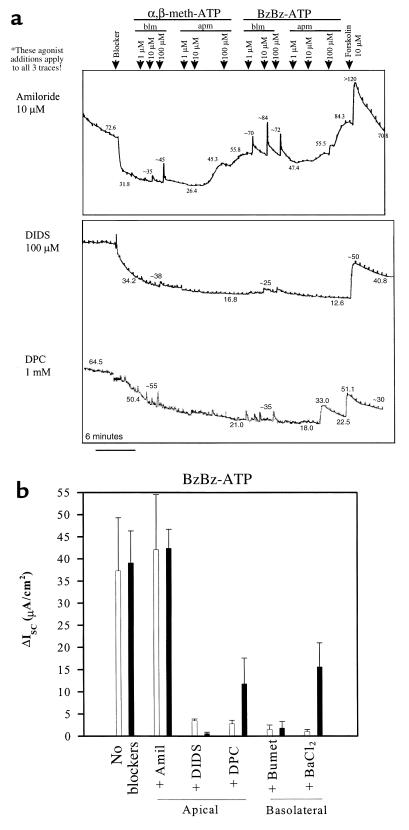
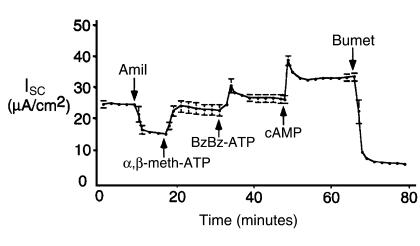
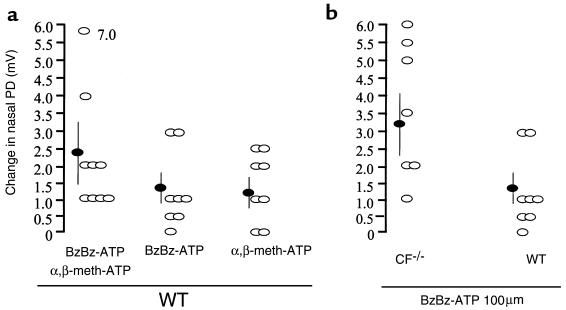

References
-
- Abbracchio MP, Burnstock G. Purinoceptors: are there families of P2X and P2Y purinoceptors? Pharmacol Ther. 1994;64:445–475. - PubMed
-
- Barnard EA, Burnstock G, Webb TE. G protein-coupled receptors for ATP and other nucleotides: a new receptor family. Trends Pharmacol Sci. 1994;15:67–70. - PubMed
-
- Buell G, Collo G, Rassendren F. P2X receptors: an emerging channel family. Eur J Neurosci. 1996;8:2221–2228. - PubMed
-
- Bean BP. Pharmacology and electrophysiology of ATP-activated ion channels. Trends Pharmacol Sci. 1992;13:87–90. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
