

GERp95, a membrane-associated protein that belongs to a family of proteins involved in stem cell differentiation
- PMID: 10512872
- PMCID: PMC25603
- DOI: 10.1091/mbc.10.10.3357
GERp95, a membrane-associated protein that belongs to a family of proteins involved in stem cell differentiation
Abstract
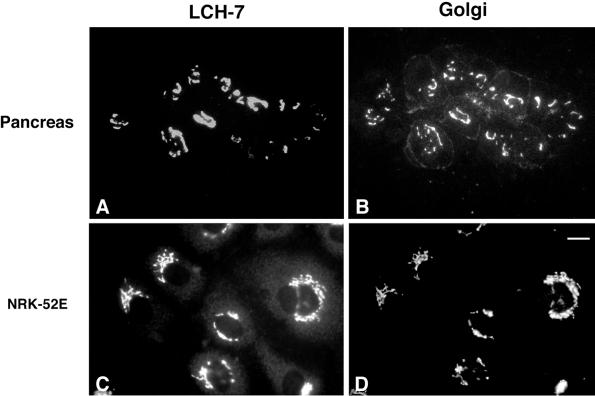
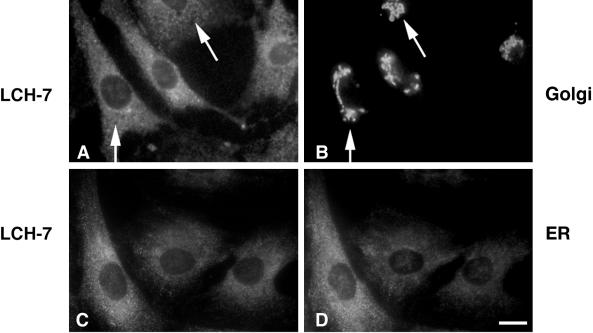
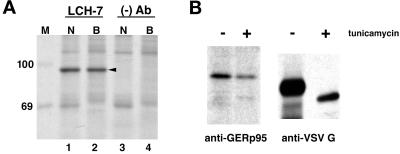
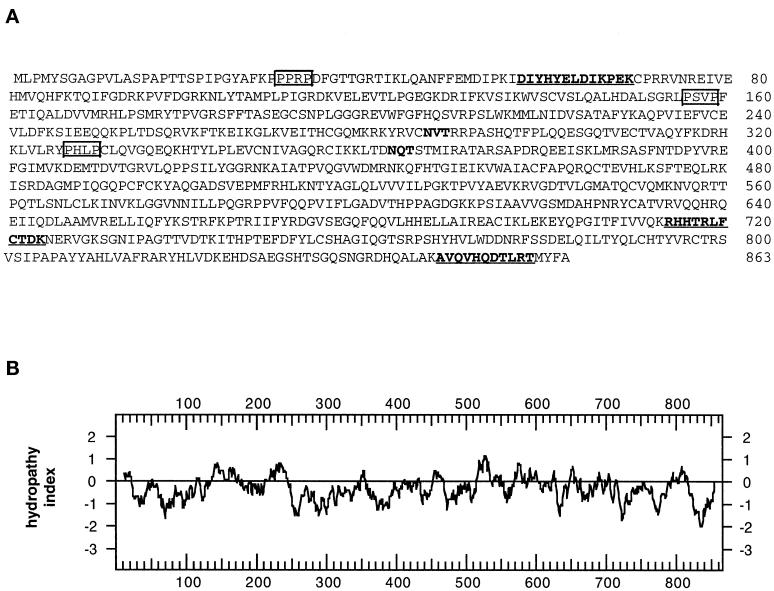
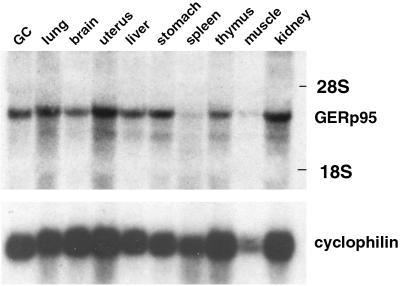
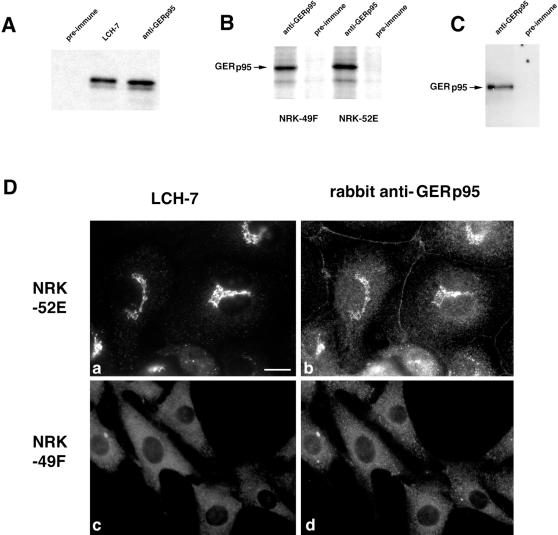
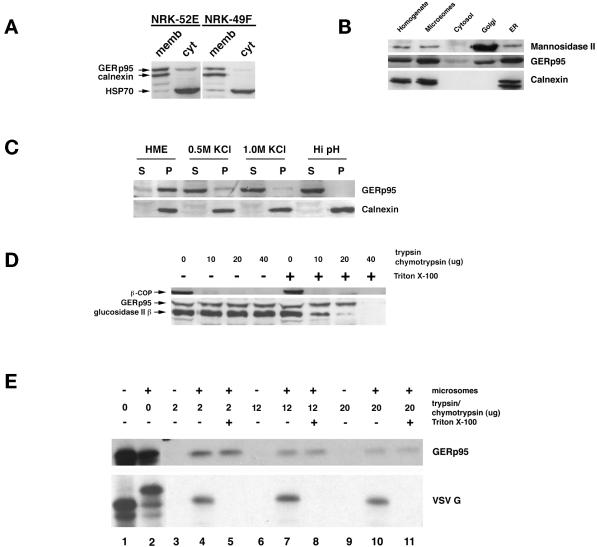
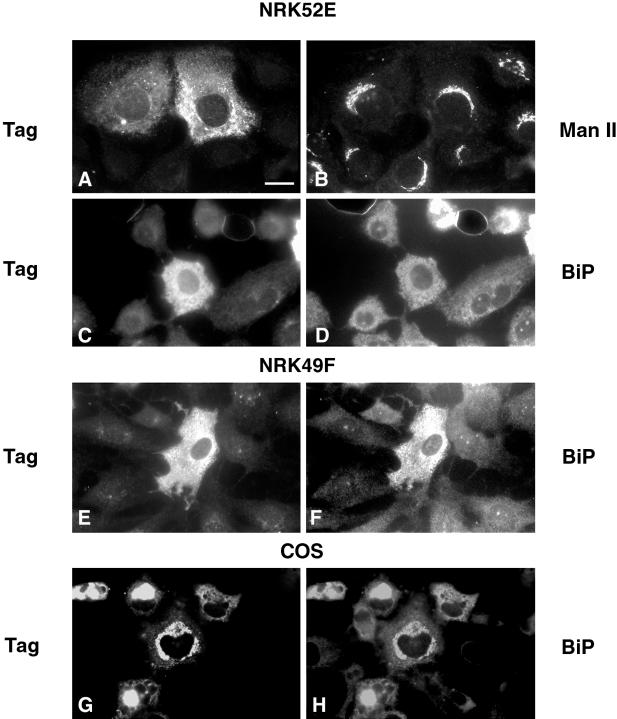
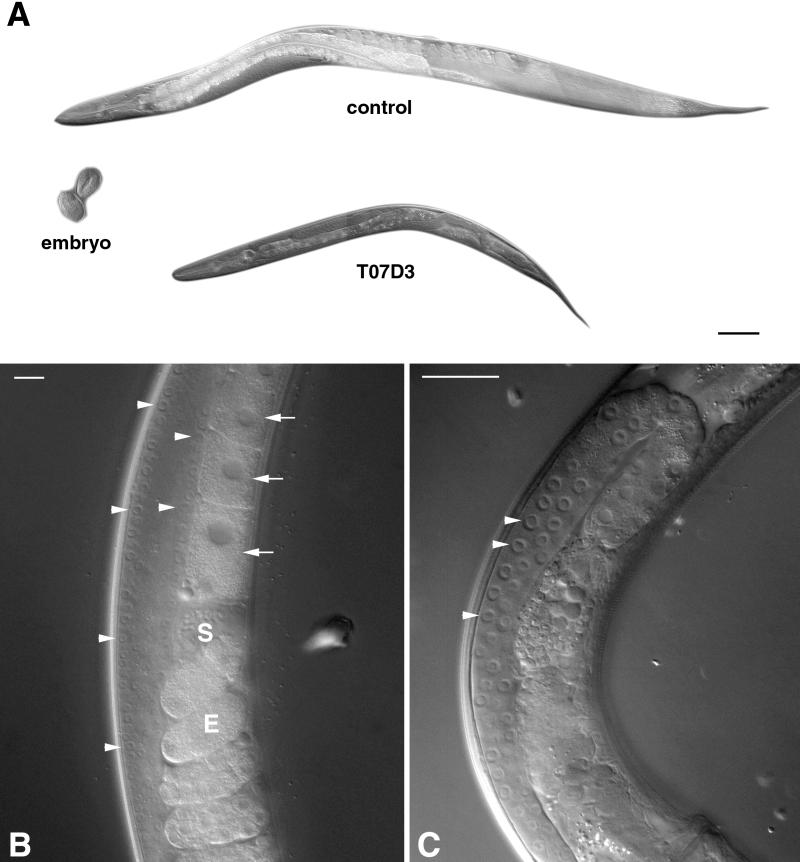
A panel of mAbs was elicited against intracellular membrane fractions from rat pancreas. One of the antibodies reacted with a 95-kDa protein that localizes primarily to the Golgi complex or the endoplasmic reticulum (ER), depending on cell type. The corresponding cDNA was cloned and sequenced and found to encode a protein of 97.6 kDa that we call GERp95 (Golgi ER protein 95 kDa). The protein copurifies with intracellular membranes but does not contain hydrophobic regions that could function as signal peptides or transmembrane domains. Biochemical analysis suggests that GERp95 is a cytoplasmically exposed peripheral membrane protein that exists in a protease-resistant complex. GERp95 belongs to a family of highly conserved proteins in metazoans and Schizosaccharomyces pombe. It has recently been determined that plant and Drosophila homologues of GERp95 are important for controlling the differentiation of stem cells (Bohmert et al., 1998; Cox et al., 1998; Moussian et al., 1998). In Caenorhabditis elegans, there are at least 20 members of this protein family. To this end, we have used RNA interference to show that the GERp95 orthologue in C. elegans is important for maturation of germ-line stem cells in the gonad. GERp95 and related proteins are an emerging new family of proteins that have important roles in metazoan development. The present study suggests that these proteins may exert their effects on cell differentiation from the level of intracellular membranes.
Figures

References
-
- Altschul S, Gish W, Miller W, Myers E, Lipman D. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Arduengo PM, Appleberry OK, Chuang P, L’Hernault SW. The presenilin protein family member SPE-4 localizes to an ER/Golgi derived organelle and is required for proper cytoplasmic partitioning during Caenorhabditis elegans spermatogenesis. J Cell Sci. 1998;111:3645–54. - PubMed
-
- Arendt CW, Ostergaard HL. Identification of the CD45-associated 116-kDa and 80-kDa proteins as the alpha- and beta-subunits of the alpha-glucosidase II. J Biol Chem. 1997;272:13117–13125. - PubMed
-
- Balch WE, Dunphy WG, Braell WA, Rothman JE. Reconstitution of the transport of protein between successive compartments of the Golgi measured by the coupled incorporation of N-acetylglucosamine. Cell. 1984;39:405–416. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
