

Genetic and biochemical characterization of a high-affinity betaine uptake system (BusA) in Lactococcus lactis reveals a new functional organization within bacterial ABC transporters
- PMID: 10515910
- PMCID: PMC103755
- DOI: 10.1128/JB.181.20.6238-6246.1999
Genetic and biochemical characterization of a high-affinity betaine uptake system (BusA) in Lactococcus lactis reveals a new functional organization within bacterial ABC transporters
Abstract
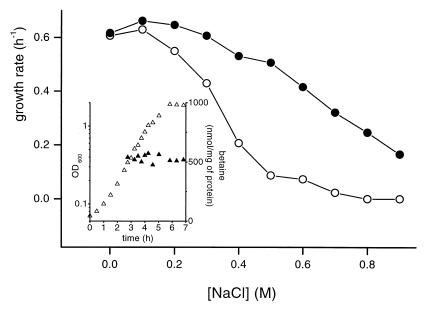
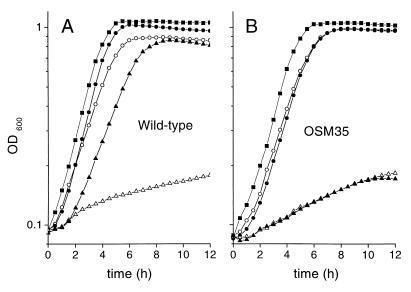
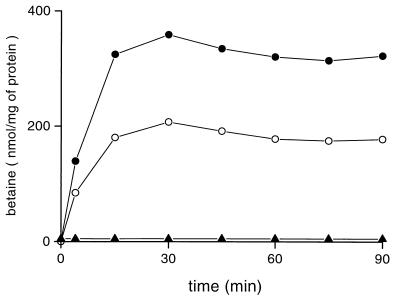
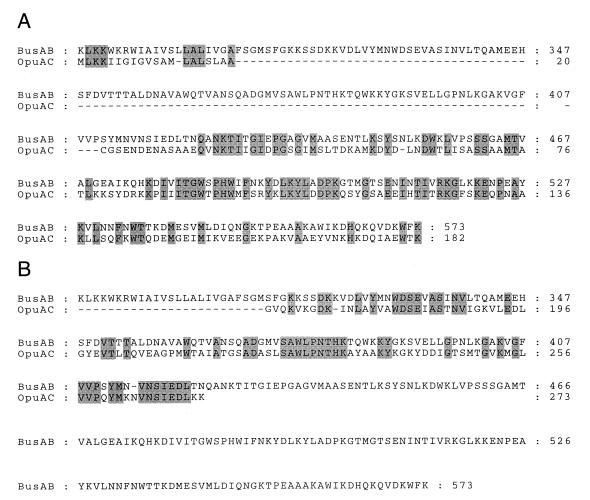
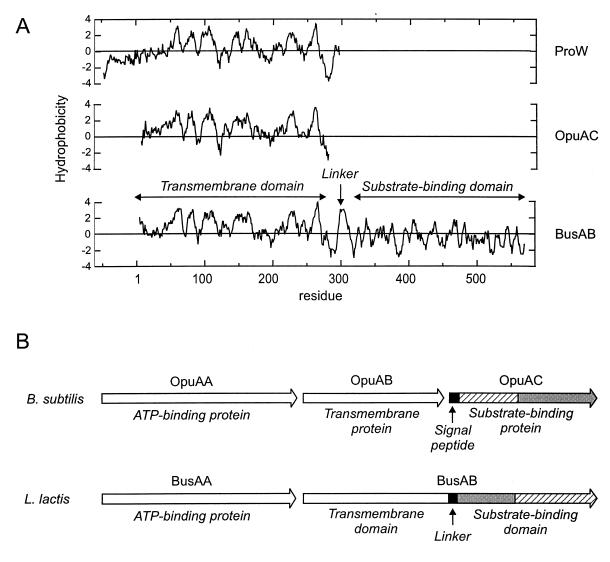
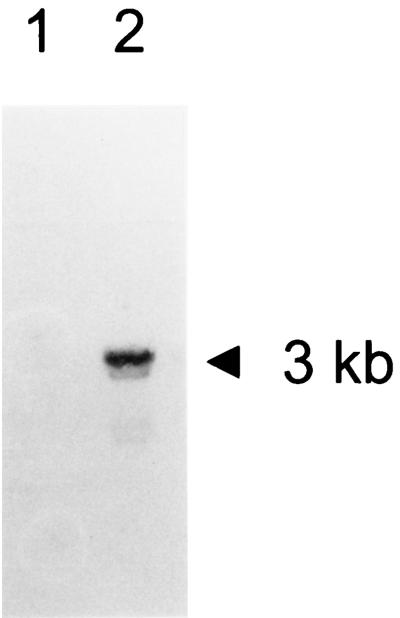
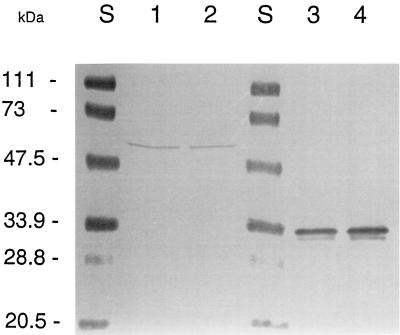
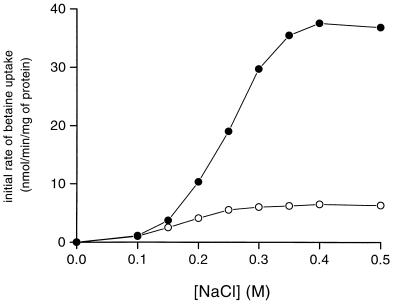
The cytoplasmic accumulation of exogenous betaine stimulates the growth of Lactococcus lactis cultivated under hyperosmotic conditions. We report that L. lactis possesses a single betaine transport system that belongs to the ATP-binding cassette (ABC) superfamily of transporters. Through transposon mutagenesis, a mutant deficient in betaine transport was isolated. We identified two genes, busAA and busAB, grouped in an operon, busA (betaine uptake system). The transcription of busA is strongly regulated by the external osmolality of the medium. The busAA gene codes for the ATP-binding protein. busAB encodes a 573-residue polypeptide which presents two striking features: (i) a fusion between the regions encoding the transmembrane domain (TMD) and the substrate-binding domain (SBD) and (ii) a swapping of the SBD subdomains when compared to the Bacillus subtilis betaine-binding protein, OpuAC. BusA of L. lactis displays a high affinity towards betaine (K(m) = 1.7 microM) and is an osmosensor whose activity is tightly regulated by external osmolality, leading the betaine uptake capacity of L. lactis to be under dual control at the biochemical and genetic levels. A protein presenting the characteristics predicted for BusAB was detected in the membrane fraction of L. lactis. The fusion between the TMD and the SBD is the first example of a new organization within prokaryotic ABC transporters.
Figures
References
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Bolotin A, Mauger S, Malarme K, Sorokin A, Ehrlich S D. Abstracts of the Conference on Streptococcal Genetics of the American Society for Microbiology, Vichy 1998. Washington, D.C.: American Society for Microbiology; 1998. Lactococcus lactis IL1403 diagnostic genomics, abstr. O3; pp. 10–11.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
