

Mutations in the DG loop of adenovirus type 5 fiber knob protein abolish high-affinity binding to its cellular receptor CAR
- PMID: 10516059
- PMCID: PMC112985
- DOI: 10.1128/JVI.73.11.9508-9514.1999
Mutations in the DG loop of adenovirus type 5 fiber knob protein abolish high-affinity binding to its cellular receptor CAR
Abstract
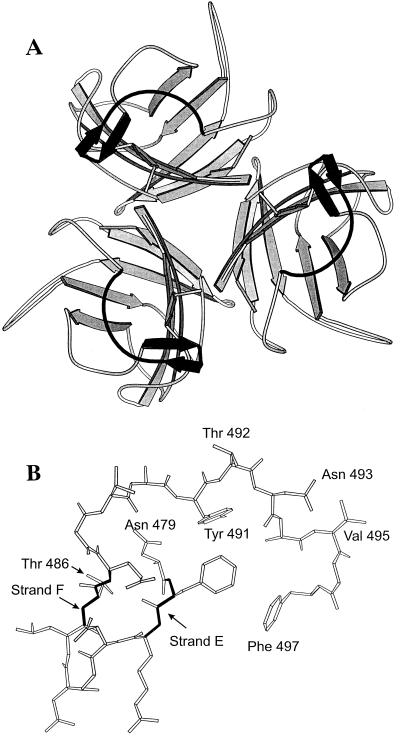
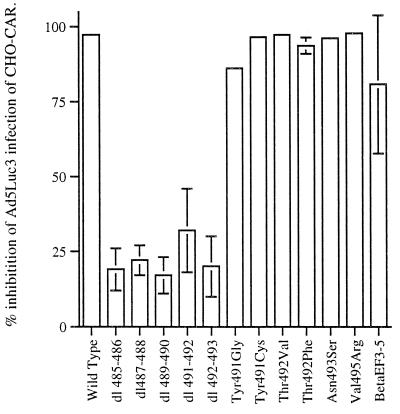
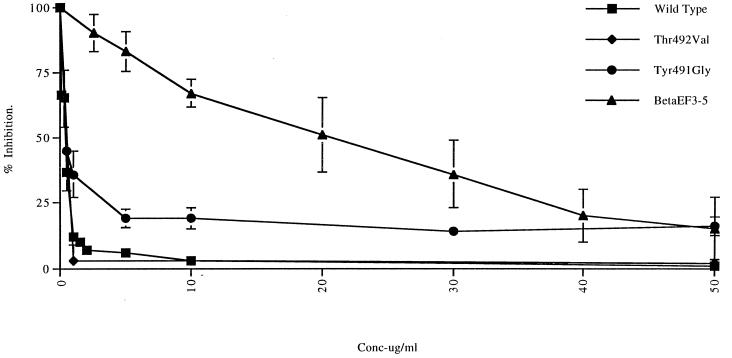
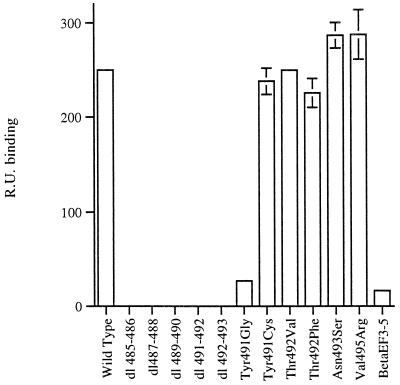
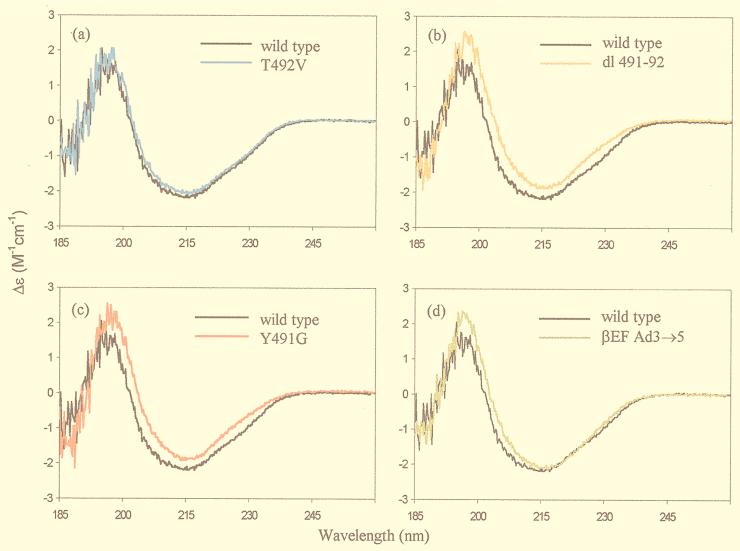
The amino acid residues in adenovirus type 5 (Ad5) fiber that interact with its cellular receptor, the coxsackie B virus and Ad receptor (CAR), have not been defined. To investigate this, multiple mutations were constructed in the region between residues 479 and 497 in Ad5 fiber (beta-strands E and F and the adjacent region of the DG loop). The effects of these mutations on binding to CAR were determined by use of cell-binding competition experiments, surface plasmon resonance, and direct binding studies. The mutation effects on the overall folding and secondary structure of the protein were assessed by circular dichroism (CD) spectroscopy. Deletions of two consecutive amino acids between residues 485 and 493 abolished high-affinity binding to CAR; the CD spectra indicated that although there was no disruption of the overall folding and secondary structure of the protein, local conformational changes did occur. Moreover, single site mutations in this region of residues with exposed, surface-accessible side chains, such as Thr492, Asn493, and Val495, had no effect on receptor binding, which demonstrates that these residues are not in contact with CAR themselves. This implies the involvement of residues in neighboring loop regions. Replacement of the segment containing the two very short beta-strands E and F and the turn between them (residues 479 to 486) with the corresponding sequence from Ad3 (betaEFAd3-->5 mutation) resulted in the loss of receptor binding. The identical CD spectra for betaEFAd3-->5 and wild-type proteins suggest that these substitutions caused no conformational rearrangement and that the loss of binding may thus be due to the substitution of one or more critical contact residues. These findings have implications for our understanding of the interaction of Ad5 fiber with CAR and for the construction of targeted recombinant Ad5 vectors for gene therapy purposes.
Figures
Similar articles
-
Identification of contact residues and definition of the CAR-binding site of adenovirus type 5 fiber protein.J Virol. 2000 Mar;74(6):2804-13. doi: 10.1128/jvi.74.6.2804-2813.2000. J Virol. 2000. PMID: 10684297 Free PMC article.
-
Adenovirus type 9 fiber knob binds to the coxsackie B virus-adenovirus receptor (CAR) with lower affinity than fiber knobs of other CAR-binding adenovirus serotypes.J Virol. 2001 Aug;75(15):7210-4. doi: 10.1128/JVI.75.15.7210-7214.2001. J Virol. 2001. PMID: 11435605 Free PMC article.
-
Adenovirus serotype 30 fiber does not mediate transduction via the coxsackie-adenovirus receptor.J Virol. 2002 Jan;76(2):656-61. doi: 10.1128/jvi.76.2.656-661.2002. J Virol. 2002. PMID: 11752156 Free PMC article.
-
Adenovirus interaction with its cellular receptor CAR.Curr Top Microbiol Immunol. 2003;272:331-64. doi: 10.1007/978-3-662-05597-7_11. Curr Top Microbiol Immunol. 2003. PMID: 12747555 Review.
-
Tropism and transduction of oncolytic adenovirus 5 vectors in cancer therapy: Focus on fiber chimerism and mosaicism, hexon and pIX.Virus Res. 2018 Sep 15;257:40-51. doi: 10.1016/j.virusres.2018.08.012. Epub 2018 Aug 17. Virus Res. 2018. PMID: 30125593 Review.
Cited by
-
A tropism-transformed Oncolytic Adenovirus with Dual Capsid Modifications for enhanced Glioblastoma Therapy.J Cancer. 2020 Jul 29;11(19):5713-5726. doi: 10.7150/jca.46463. eCollection 2020. J Cancer. 2020. PMID: 32913465 Free PMC article.
-
Mechanism of ad5 vaccine immunity and toxicity: fiber shaft targeting of dendritic cells.PLoS Pathog. 2007 Feb;3(2):e25. doi: 10.1371/journal.ppat.0030025. PLoS Pathog. 2007. PMID: 17319743 Free PMC article.
-
CAR Co-Operates With Integrins to Promote Lung Cancer Cell Adhesion and Invasion.Front Oncol. 2022 Feb 14;12:829313. doi: 10.3389/fonc.2022.829313. eCollection 2022. Front Oncol. 2022. PMID: 35252000 Free PMC article.
-
Genetic retargeting of adenovirus: novel strategy employing "deknobbing" of the fiber.J Virol. 2001 Aug;75(16):7280-9. doi: 10.1128/JVI.75.16.7280-7289.2001. J Virol. 2001. PMID: 11462000 Free PMC article.
-
Adenoviral gene therapy, radiation, and prostate cancer.Rev Urol. 2005 Fall;7(4):193-202. Rev Urol. 2005. PMID: 16985830 Free PMC article.
References
-
- Belin M, Boulanger P A. Involvement of cellular adhesion sequences in the attachment of adenovirus to the HeLa cell surface. J Gen Virol. 1994;74:1485–1497. - PubMed
-
- Bergelson J M, Cunningham J A, Droguett G, Kurt-Jones E A, Krithivas A, Hong J, Horwitz M S, Crowell R L, Finberg R W. Isolation of a common receptor for coxsackie B viruses and adenoviruses 2 and 5. Science. 1997;275:1320–1323. - PubMed
-
- Cook J P D, Henry A J, McDonnell J M, Owens R J, Sutton B J, Gould H J. Identification of contact residues in the IgE binding sites of human FceRIa. Biochemistry. 1997;36:15579–15588. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
