

Temporal and spatial analysis of Sin Nombre virus quasispecies in naturally infected rodents
- PMID: 10516063
- PMCID: PMC112989
- DOI: 10.1128/JVI.73.11.9544-9554.1999
Temporal and spatial analysis of Sin Nombre virus quasispecies in naturally infected rodents
Abstract
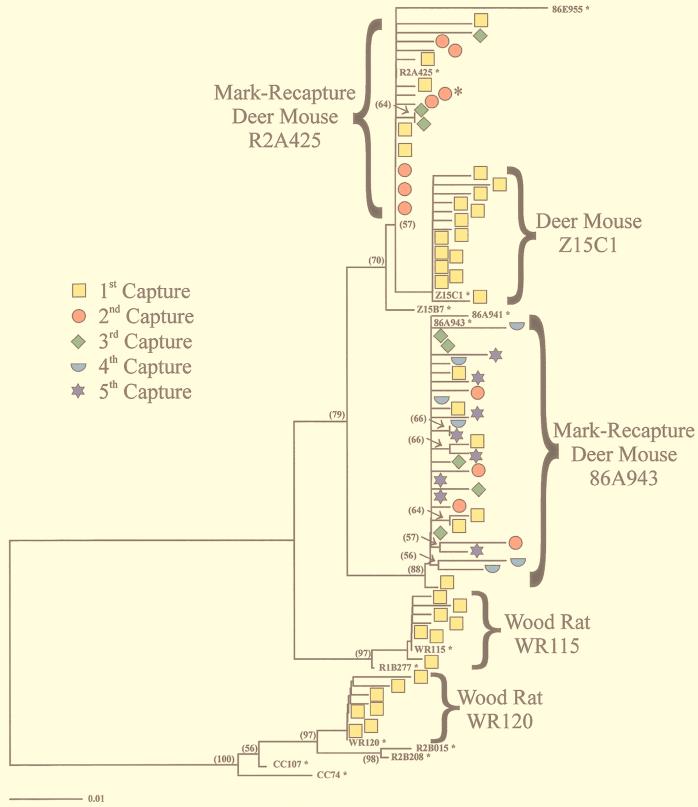
Sin Nombre virus (SNV) is thought to establish a persistent infection in its natural reservoir, the deer mouse (Peromyscus maniculatus), despite a strong host immune response. SNV-specific neutralizing antibodies were routinely detected in deer mice which maintained virus RNA in the blood and lungs. To determine whether viral diversity played a role in SNV persistence and immune escape in deer mice, we measured the prevalence of virus quasispecies in infected rodents over time in a natural setting. Mark-recapture studies provided serial blood samples from naturally infected deer mice, which were sequentially analyzed for SNV diversity. Viral RNA was detected over a period of months in these rodents in the presence of circulating antibodies specific for SNV. Nucleotide and amino acid substitutions were observed in viral clones from all time points analyzed, including changes in the immunodominant domain of glycoprotein 1 and the 3' small segment noncoding region of the genome. Viral RNA was also detected in seven different organs of sacrificed deer mice. Analysis of organ-specific viral clones revealed major disparities in the level of viral diversity between organs, specifically between the spleen (high diversity) and the lung and liver (low diversity). These results demonstrate the ability of SNV to mutate and generate quasispecies in vivo, which may have implications for viral persistence and possible escape from the host immune system.
Figures
References
-
- Abbas A K, Lichtman A H, Pober J S. Cellular and molecular immunology. Philadelphia, Pa: W. B. Saunders Company; 1998. pp. 14–31.
-
- Bharadwaj M, Botten J, Torrez-Martinez N, Hjelle B. Rio Mamore virus: genetic characterization of a newly recognized hantavirus of the pygmy rice rat, Oligoryzomys microtis, from Bolivia. Am J Trop Med Hyg. 1997;57:368–374. - PubMed
-
- Boone J D, Otteson E W, McGwire K C, Villard P, Rowe J E, St. Jeor S C. Ecology and demographics of hantavirus infections in rodent populations in the Walker River Basin of Nevada and California. Am J Trop Med Hyg. 1998;59:445–451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
