

Rapid repression of maize invertases by low oxygen. Invertase/sucrose synthase balance, sugar signaling potential, and seedling survival
- PMID: 10517852
- PMCID: PMC59423
- DOI: 10.1104/pp.121.2.599
Rapid repression of maize invertases by low oxygen. Invertase/sucrose synthase balance, sugar signaling potential, and seedling survival
Abstract
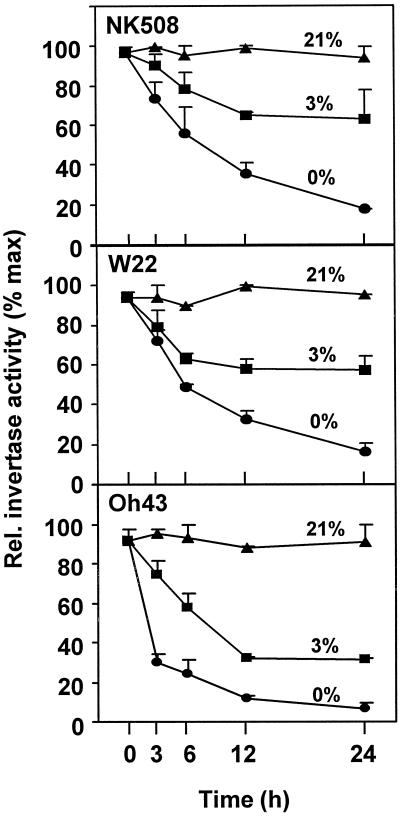
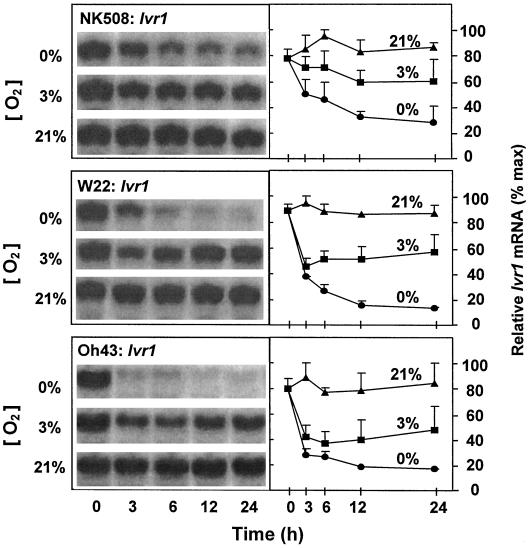
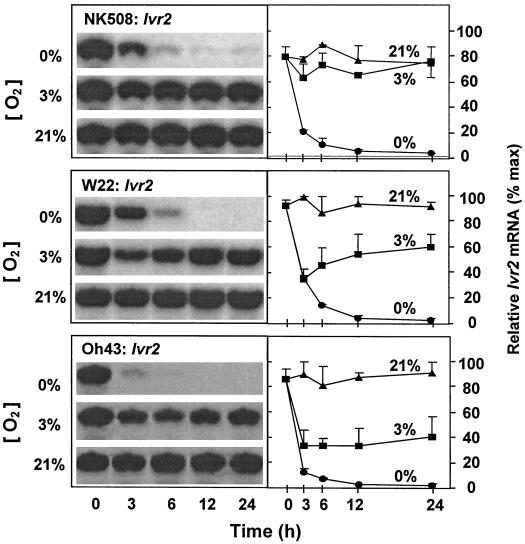
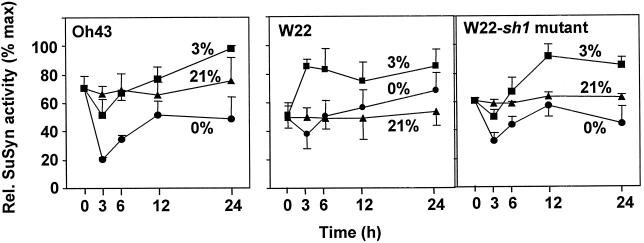
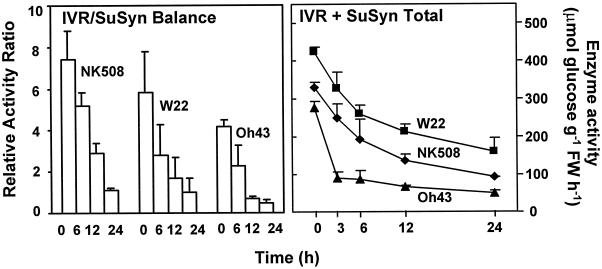
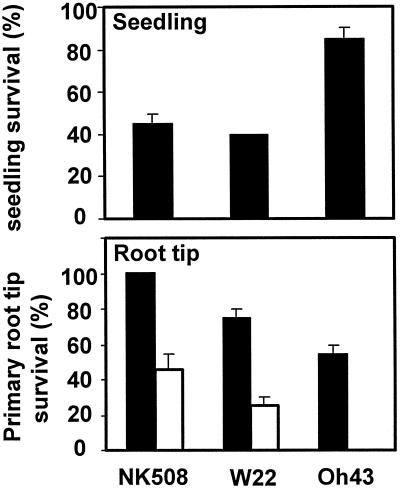
We show here that invertase gene expression and the invertase-sucrose (Suc) synthase ratio decrease abruptly in response to low oxygen in maize root tips. In addition to aiding in the conservation of carbon and possibly ATP, this response has the potential to directly affect sugar signaling relative to carbon flux. Experiments were motivated by the potential for a reduced invertase/Suc synthase balance to alter the impact of respiratory and/or membrane carbon flux on sugar signaling. Maize (Zea mays L.) seedlings with 5-cm primary roots were exposed to anoxic (0% [v/v] O2), hypoxic (3% [v/v] O2), and aerobic conditions. Rapid repression of the Ivr1 and Ivr2 maize invertases by low oxygen was evident in root tips within 3 h at both the transcript and activity levels. The speed and extent of this response increased with the degree of oxygen deprivation and differed with genotypes. This decrease in expression also contrasted markedly to that of other genes for respiratory Suc metabolism, such as Suc synthases, which typically increased or remained constant. Although previous work showed that the contrasting effects of sugars on Suc synthase genes were reflected in their regulation by hypoxia and anoxia, the same was not observed for the differentially sugar-responsive invertases. Theoretically advantageous reductions in the invertase/Suc synthase balance thus resulted. However, where this response was extreme (an Oh43 inbred), total sucrolytic capacity dropped below an apparent minimum and root tip viability was reduced. Paradoxically, only Oh43 seedlings showed survival levels >80% (versus <50%) after extreme, long-term stress, suggesting a possible advantage for multiple means of reducing sink activity. Overall, our results demonstrate a rapid change in the regulation and balance of invertases and Suc synthases that could have an immediate impact on limiting the extent of Suc cleavage and reducing the extent of concomitant, hexose-based sugar signaling under low oxygen.
Figures
References
-
- Bailey-Serres J, Kloeckner-Gruissem B, Freeling M. Genetic and molecular approaches to the study of the anaerobic responses and tissue specific gene expression in maize. Plant Cell Environ. 1988;11:351–357.
-
- Bell GI, Pilkis SJ, Weber IT, Polonsky KS. Glucokinase mutations, insulin secretion, and Diabetes mellitus. Annu Rev Physiol. 1996;58:171–186. - PubMed
-
- Bertani A, Brambilla I, Menegus F. Effect of anaerobiosis on carbohydrate content in rice roots. Biochem Physiol Pflanz. 1981;176:835–840.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
