

Rates of sugar uptake by guard cell protoplasts of pisum sativum L. Related To the solute requirement for stomatal opening
- PMID: 10517857
- PMCID: PMC59428
- DOI: 10.1104/pp.121.2.647
Rates of sugar uptake by guard cell protoplasts of pisum sativum L. Related To the solute requirement for stomatal opening
Abstract
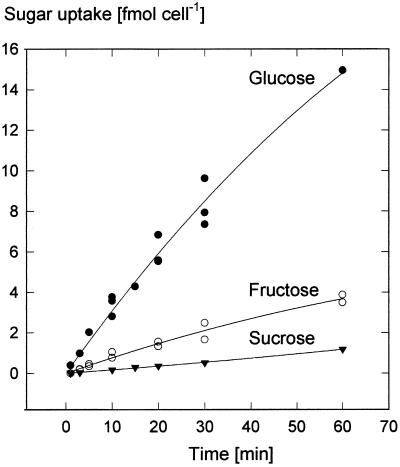
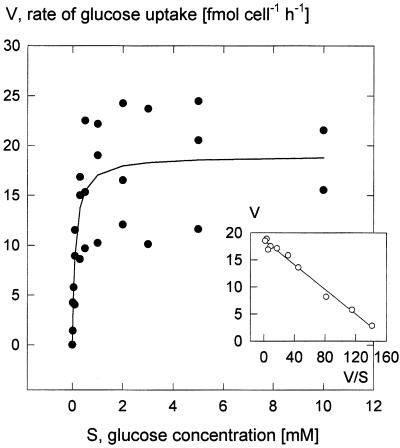
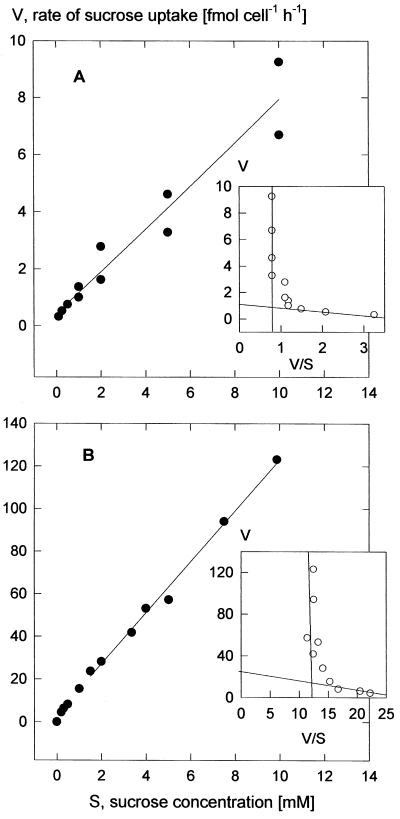
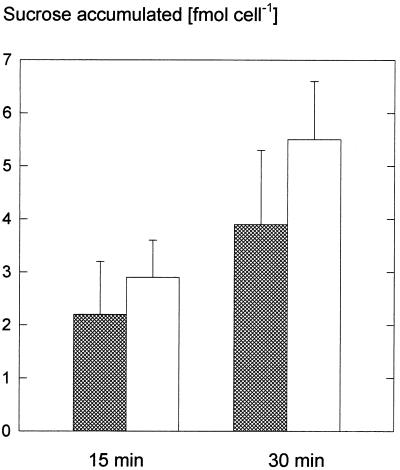
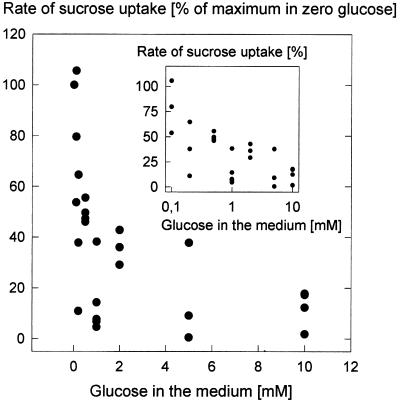
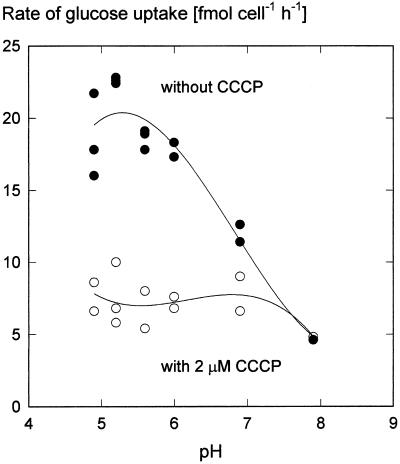
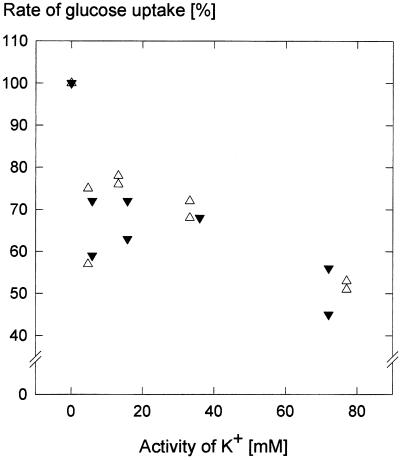
We wished to determine whether the capacity of the sugar uptake mechanisms of guard cells of the Argenteum mutant of pea (Pisum sativum L.) sufficed to support a concurrent stomatal opening movement. Sugar uptake by guard cell protoplasts was determined by silicone-oil-filtering centrifugation. The protoplasts took up [(14)C]glucose, [(14)C]fructose, and [(14)C]sucrose (Suc), apparently in symport with protons. Mannose, galactose, and fructose competed with Glc for transport by a presumed hexose carrier. The uptake of Glc saturated with a K(m) of 0.12 mM and a V(max) of 19 fmol cell(-1) h(-1). At external concentrations <1 mM, the uptake of Suc was slower than that of Glc. It exhibited a saturating component with a K(m) varying between 0.25 and 0.8 mM and a V(max) between 1 and 10 fmol cell(-1) h(-1), and at external concentrations >1 mM, a non-saturating component. At apoplastic sugar concentrations below 4 mM, sugar import was estimated to be mainly in the form of hexoses and too slow to support a simultaneous stomatal opening movement. If, however, during times of high photosynthesis and transpiration, the apoplastic Suc concentration rose and entered the range of non-saturating import, absorbed Suc could replace potassium malate as the osmoticum for the maintenance of stomatal opening.
Figures
References
-
- Aked J, Hall JL. The uptake of glucose, fructose and sucrose into the lower epidermis of leaf discs of pea (Pisum sativum L. cv. Argenteum) New Phytol. 1993;123:271–276.
-
- Boorer KJ, Loo DDF, Frommer WB, Wright EM. Transport mechanism of the cloned potato H+-sucrose cotransporter StSUT1. J Biol Chem. 1996;271:25139–25144. - PubMed
-
- Borchert S, Grosse H, Heldt HW. Specific transport of inorganic phosphate, dihydroxyacetone phosphate and 3-phosphoglycerate into amyloplasts from pea roots. FEBS Lett. 1989;253:183–186.
-
- Buckhout TJ. Sucrose transport in isolated plasma-membrane vesicles from sugar beet (Beta vulgaris L.): evidence for an electrogenic sucrose-proton symport. Planta. 1989;178:393–399. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
