

Analysis of the relative increase in photosynthetic O(2) uptake when photosynthesis in grapevine leaves is inhibited following low night temperatures and/or water stress
- PMID: 10517860
- PMCID: PMC59431
- DOI: 10.1104/pp.121.2.675
Analysis of the relative increase in photosynthetic O(2) uptake when photosynthesis in grapevine leaves is inhibited following low night temperatures and/or water stress
Abstract
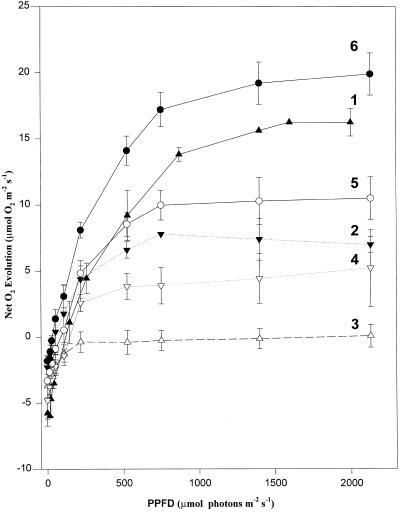
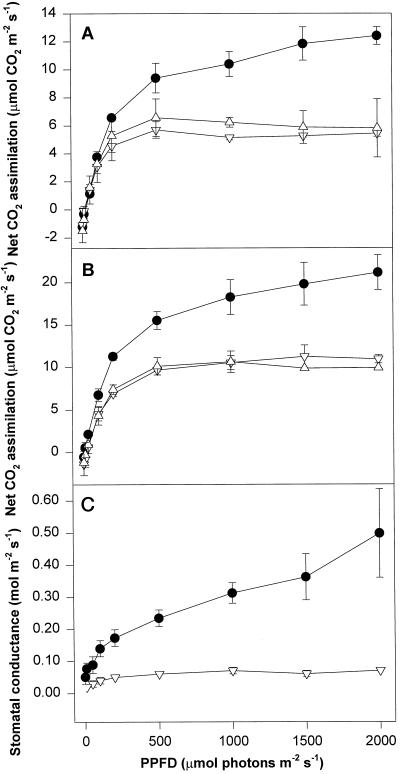
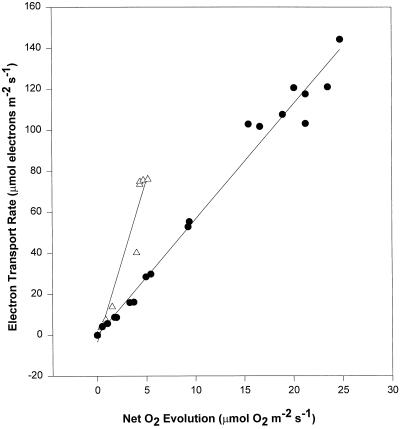
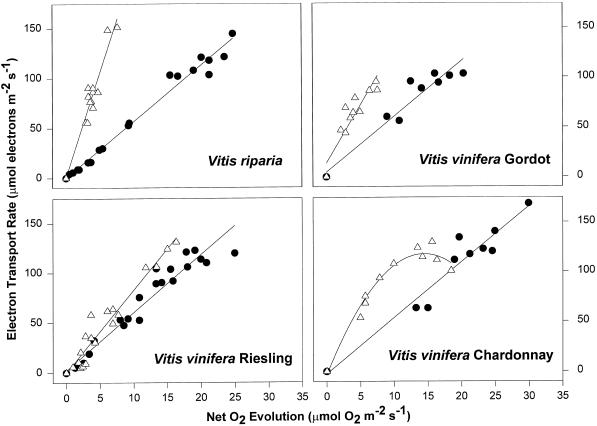
We found similarities between the effects of low night temperatures (5 degrees C-10 degrees C) and slowly imposed water stress on photosynthesis in grapevine (Vitis vinifera L.) leaves. Exposure of plants growing outdoors to successive chilling nights caused light- and CO(2)-saturated photosynthetic O(2) evolution to decline to zero within 5 d. Plants recovered after four warm nights. These photosynthetic responses were confirmed in potted plants, even when roots were heated. The inhibitory effects of chilling were greater after a period of illumination, probably because transpiration induced higher water deficit. Stomatal closure only accounted for part of the inhibition of photosynthesis. Fluorescence measurements showed no evidence of photoinhibition, but nonphotochemical quenching increased in stressed plants. The most characteristic response to both stresses was an increase in the ratio of electron transport to net O(2) evolution, even at high external CO(2) concentrations. Oxygen isotope exchange revealed that this imbalance was due to increased O(2) uptake, which probably has two components: photorespiration and the Mehler reaction. Chilling- and drought-induced water stress enhanced both O(2) uptake processes, and both processes maintained relatively high rates of electron flow as CO(2) exchange approached zero in stressed leaves. Presumably, high electron transport associated with O(2) uptake processes also maintained a high DeltapH, thus affording photoprotection.
Figures

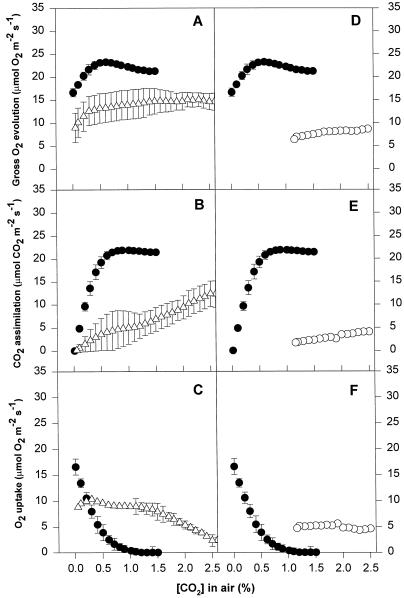
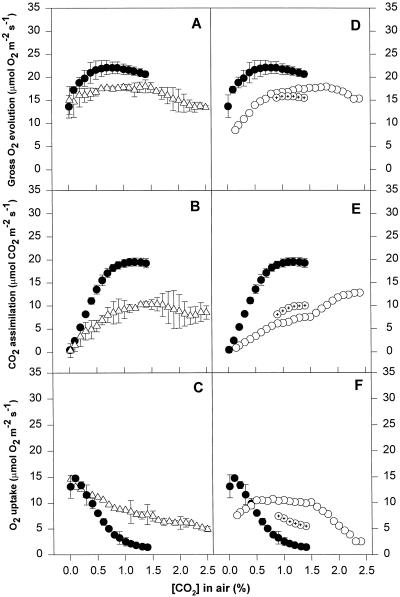
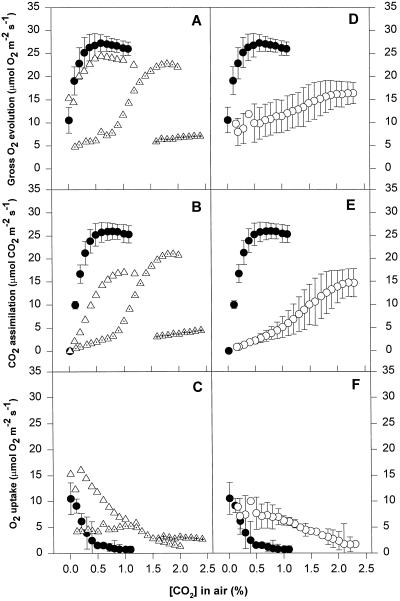
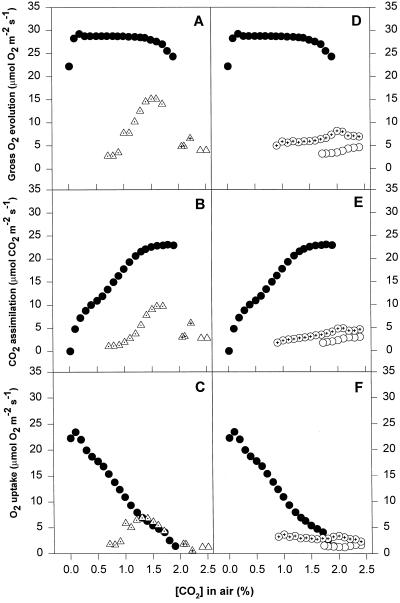
References
-
- Asada K, Nakano Y. Affinity for oxygen in photoreduction of molecular oxygen and scavenging of hydrogen peroxide in spinach chloroplasts. Photochem Photobiol. 1978;28:917–920.
-
- Badger MR. Photosynthetic oxygen exchange. Annu Rev Plant Physiol. 1985;36:27–53.
-
- Báló B, Lajkó F, Garab G. Effects of chilling on photosynthesis of grapevines. Photosynthetica. 1991;25:227–230.
-
- Berry J, Björkman O. Photosynthetic response and adaptation to temperature in higher plants. Annu Rev Plant Physiol. 1980;31:491–543.
LinkOut - more resources
Full Text Sources
