

Imino sugars inhibit the formation and secretion of bovine viral diarrhea virus, a pestivirus model of hepatitis C virus: implications for the development of broad spectrum anti-hepatitis virus agents
- PMID: 10518544
- PMCID: PMC18380
- DOI: 10.1073/pnas.96.21.11878
Imino sugars inhibit the formation and secretion of bovine viral diarrhea virus, a pestivirus model of hepatitis C virus: implications for the development of broad spectrum anti-hepatitis virus agents
Abstract
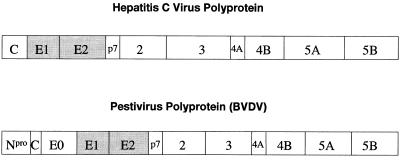
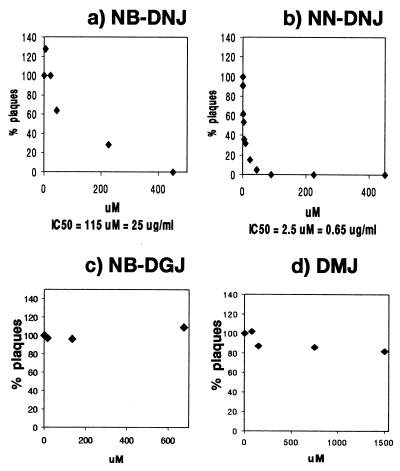
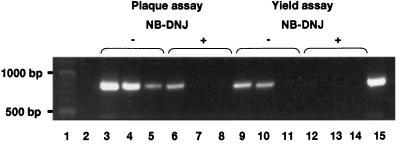
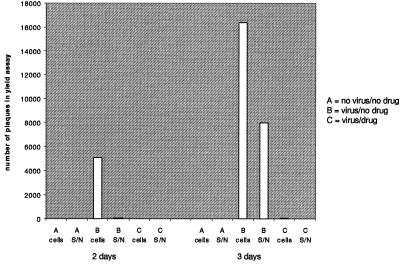
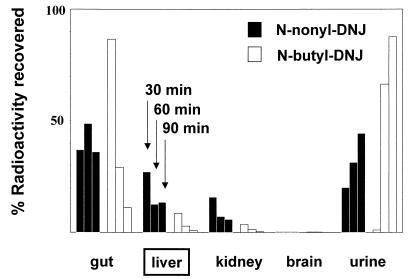
One function of N-linked glycans is to assist in the folding of glycoproteins by mediating interactions of the lectin-like chaperone proteins calnexin and calreticulin with nascent glycoproteins. These interactions can be prevented by inhibitors of the alpha-glucosidases, such as N-butyl-deoxynojirimycin (NB-DNJ) and N-nonyl-DNJ (NN-DNJ), and this causes some proteins to be misfolded and retained within the endoplasmic reticulum (ER). We have shown previously that the NN-DNJ-induced misfolding of one of the hepatitis B virus (HBV) envelope glycoproteins prevents the formation and secretion of virus in vitro and that this inhibitor alters glycosylation and reduces the viral levels in an animal model of chronic HBV infection. This led us to investigate the effect of glucosidase inhibitors on another ER-budding virus, bovine viral diarrhea virus, a tissue culture surrogate of human hepatitis C virus (HCV). Here we show that in MDBK cells alpha-glucosidase inhibitors prevented the formation and secretion of infectious bovine viral diarrhea virus. Data also are presented showing that NN-DNJ, compared with NB-DNJ, exhibits a prolonged retention in liver in vivo. Because viral secretion is selectively hypersensitive to glucosidase inhibition relative to the secretion of cellular proteins, the possibility that glucosidase inhibitors could be used as broad-based antiviral hepatitis agents is discussed. A single drug against HBV, HCV, and, possibly, HDV, which together chronically infect more than 400 million people worldwide, would be of great therapeutic value.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
