

Reconstitution of a minimal RNA degradosome demonstrates functional coordination between a 3' exonuclease and a DEAD-box RNA helicase
- PMID: 10521403
- PMCID: PMC317069
- DOI: 10.1101/gad.13.19.2594
Reconstitution of a minimal RNA degradosome demonstrates functional coordination between a 3' exonuclease and a DEAD-box RNA helicase
Erratum in
- Genes Dev 2000 May 1;14(9):1167
Abstract
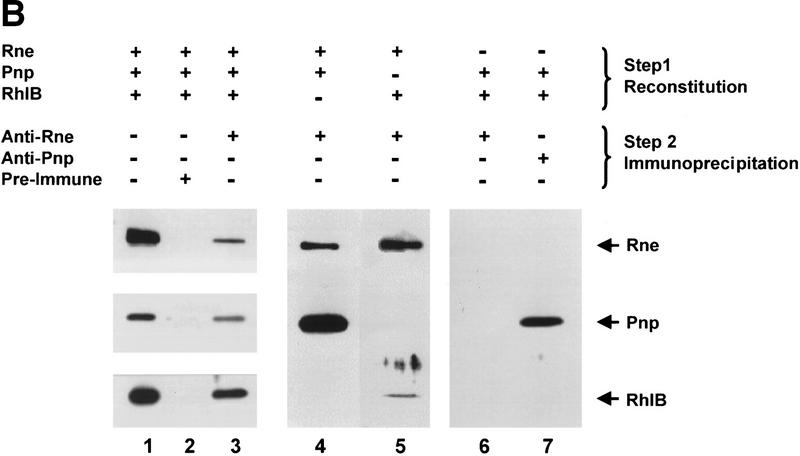
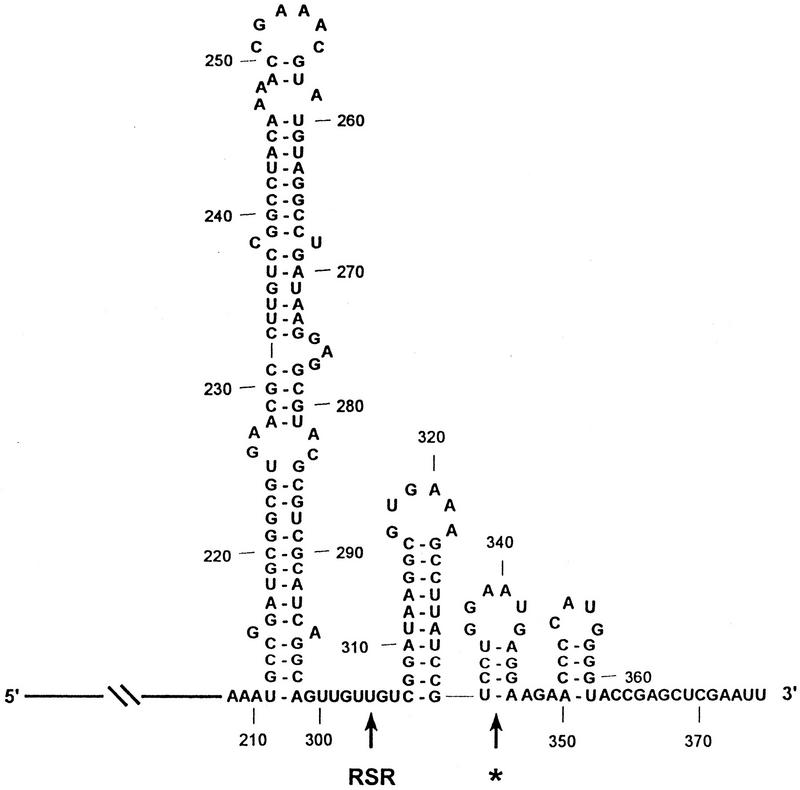
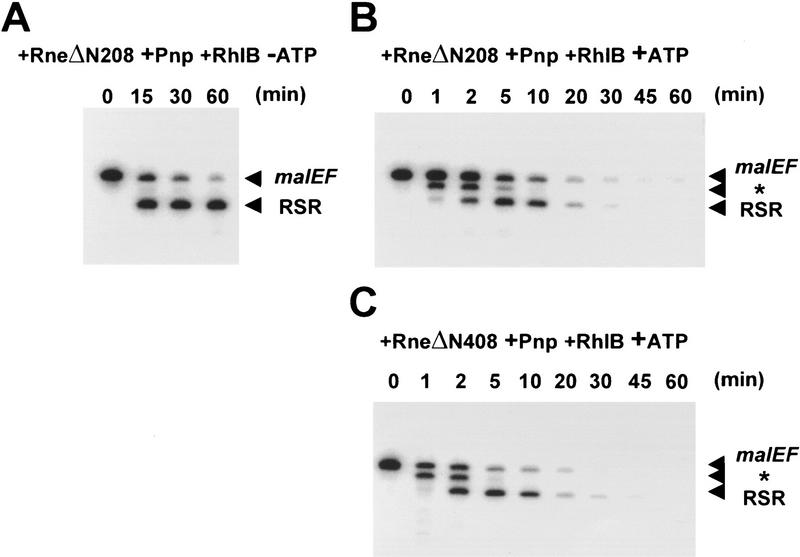
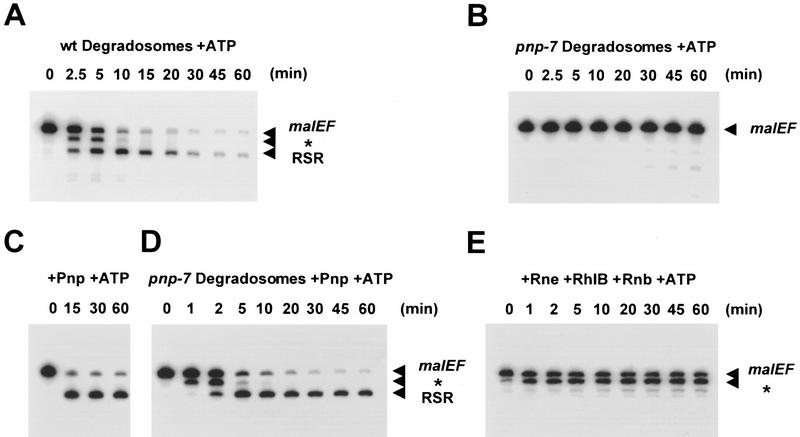
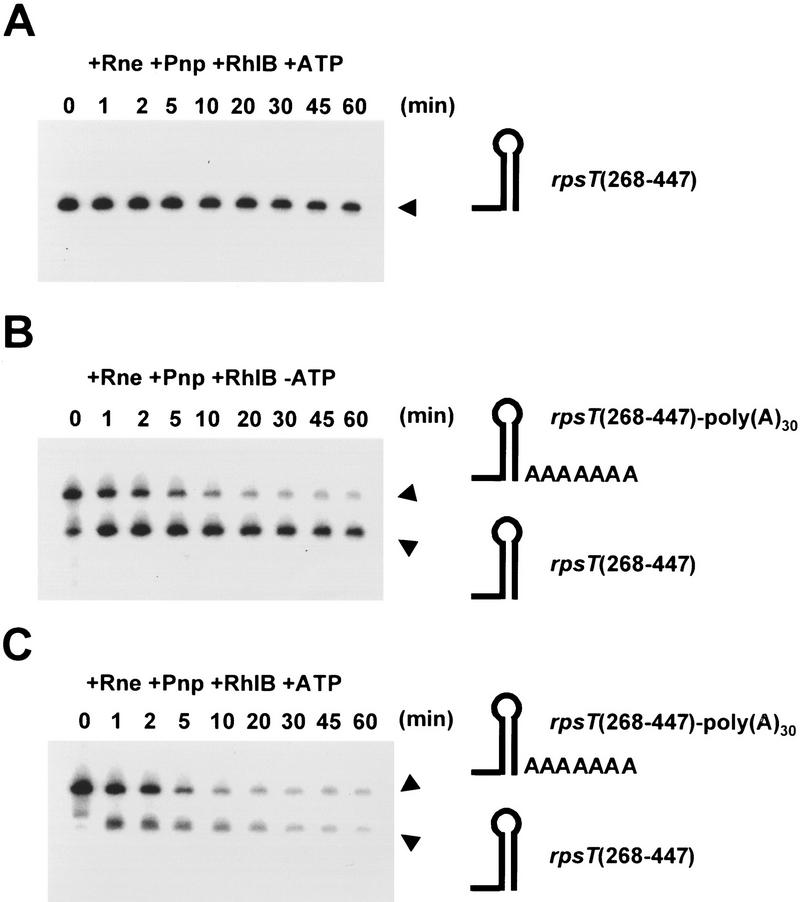
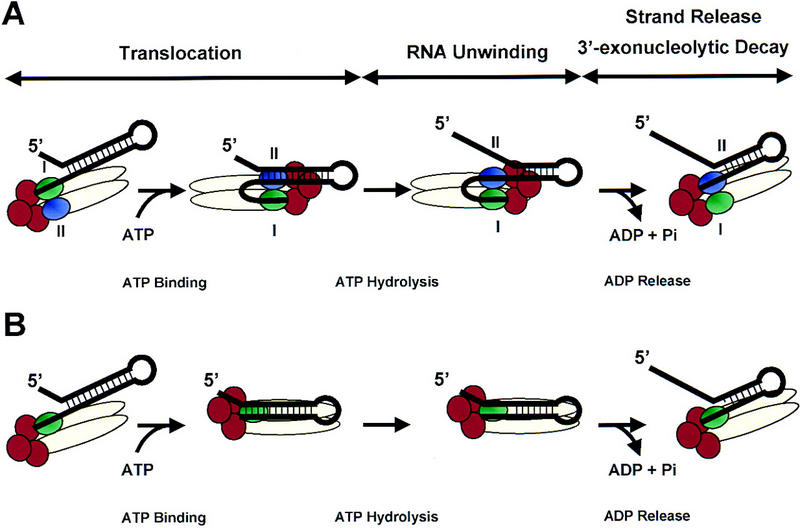
The RNA degradosome is a multiprotein complex required for the degradation of highly structured RNAs. We have developed a method for reconstituting a minimal degradosome from purified proteins. Our results demonstrate that a degradosome-like complex containing RNase E, PNPase, and RhlB can form spontaneously in vitro in the absence of all other cellular components. Moreover, ATP-dependent degradation of the malEF REP RNA by the reconstituted, minimal degradosome is indistinguishable from that of degradosomes isolated from whole cells. The Rne protein serves as an essential scaffold in the reconstitution process; however, RNase E activity is not required. Rather, Rne coordinates the activation of RhlB dependent on a 3' single-stranded extension on RNA substrates. A model for degradosome-mediated degradation of structured RNA is presented with its implications for mRNA decay in Escherichia coli.
Figures


References
-
- Alberts B. The cell as a collection of protein machines: Preparing the next generation of molecular biologists. Cell. 1998;92:291–294. - PubMed
-
- Blum E, Py B, Carpousis AJ, Higgins CF. Polyphosphate kinase is a component of the Escherichia coli RNA degradosome. Mol Microbiol. 1997;26:387–398. - PubMed
-
- Blum E, Carpousis AJ, Higgins CF. Polyadenylation promotes degradation of 3′-structured RNA by the Escherichia coli mRNA degradosome in vitro. J Biol Chem. 1999;274:4009–4016. - PubMed
-
- Carpousis AJ, Van Houwe G, Ehretsmann C, Krisch HM. Copurification of E. coli RNase E and PNPase: Evidence for a specific association between two enzymes important for RNA processing and degradation. Cell. 1994;76:889–900. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases