

Herpes simplex virus type 1 infection of activated cytotoxic T cells: Induction of fratricide as a mechanism of viral immune evasion
- PMID: 10523608
- PMCID: PMC2195666
- DOI: 10.1084/jem.190.8.1103
Herpes simplex virus type 1 infection of activated cytotoxic T cells: Induction of fratricide as a mechanism of viral immune evasion
Abstract
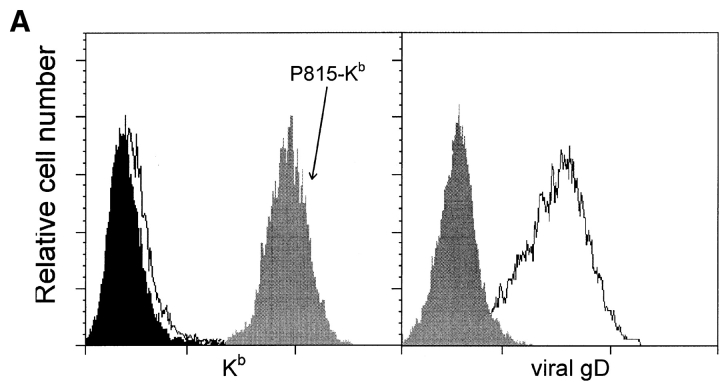
Herpes simplex virus type 1 (HSV1), a large DNA-containing virus, is endemic in all human populations investigated. After infection of mucocutaneous surfaces, HSV1 establishes a latent infection in nerve cells. Recently, it was demonstrated that HSV1 can also infect activated T lymphocytes. However, the consequences of T cell infection for viral pathogenesis and immunity are unknown. We have observed that in contrast to the situation in human fibroblasts, in human T cell lines antigen presentation by major histocompatibility complex class I molecules is not blocked after HSV1 infection. Moreover, HSV1 infection of T cells results in rapid elimination of antiviral T cells by fratricide. To dissect the underlying molecular events, we used a transgenic mouse model of HSV1 infection to demonstrate that CD95 (Apo-1, Fas)-triggered apoptosis is essential for HSV1-induced fratricide, whereas tumor necrosis factor (TNF) also contributes to this phenomenon but to a lesser extent. By contrast, neither TRAIL (TNF-related apoptosis-inducing ligand) nor perforin were involved. Finally, we defined two mechanisms associated with HSV1-associated fratricide of antiviral T cells: (a) T cell receptor-mediated upregulation of CD95 ligand and (b) a viral "competence-to-die" signal that renders activated T lymphocytes susceptible to CD95 signaling. We propose that induction of fratricide is an important immune evasion mechanism of HSV1, helping the virus to persist in the host organism throughout its lifetime.
Figures









References
-
- Roizman B., Sears A.E. Herpes simplex viruses and their replication. In: Fields B.N., Knipe D.M., Howley P.M., editors. Virology. Lippincott-Raven Publishers; Philadelphia: 1996. pp. 2231–2295.
-
- Whitley R.J. Herpes simplex viruses. In: Fields B.N., Knipe D.M., Howley P.M., editors. Virology. Lippincott-Raven Publishers; Philadelphia: 1996. pp. 2297–2342.
-
- Ward P.L., Roizman B. Evasion and obstructionthe central strategy of the interaction of human herpesviruses with host defense. In: Medveczky P.G., Friedman H., Bendinelli M., editors. Herpesviruses and Immunity. Plenum Press; New York: 1998. pp. 1–32.
-
- Rouse B.T., Atherton S.S. Immunopathology of herpesvirus infection. In: Medveczky P.G., Friedman H., Bendinelli M., editors. Herpesviruses and Immunity. Plenum Press; New York: 1998. pp. 33–51.
-
- Cose S.C., Jones C.M., Wallace M.E., Heath W.R., Carbone F.R. Antigen-specific CD8+ T cell subset distribution in lymph nodes draining the site of herpes simplex virus infection. Eur. J. Immunol. 1997;27:2310–2316. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
