

Protective T cell-independent antiviral antibody responses are dependent on complement
- PMID: 10523614
- PMCID: PMC2195668
- DOI: 10.1084/jem.190.8.1165
Protective T cell-independent antiviral antibody responses are dependent on complement
Abstract
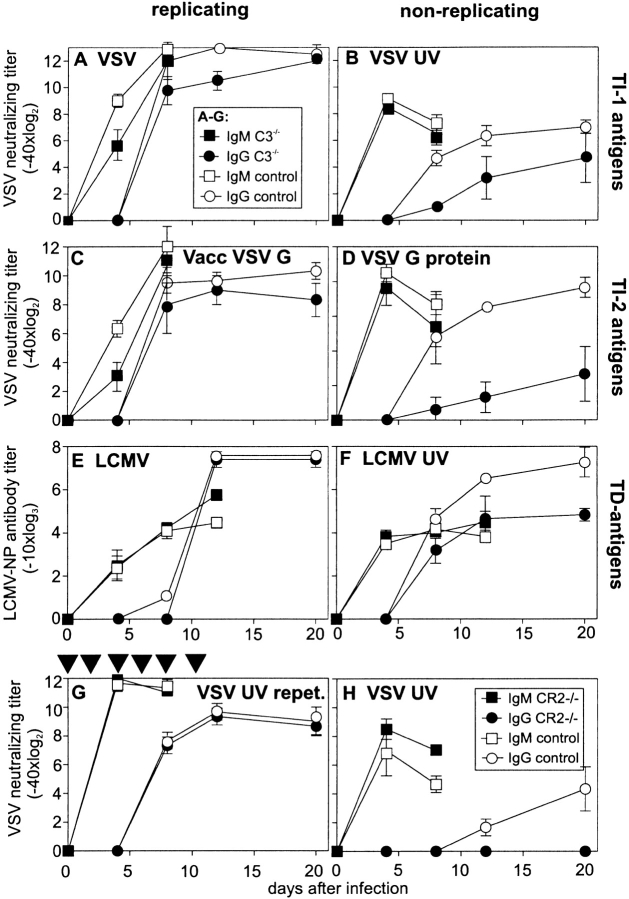
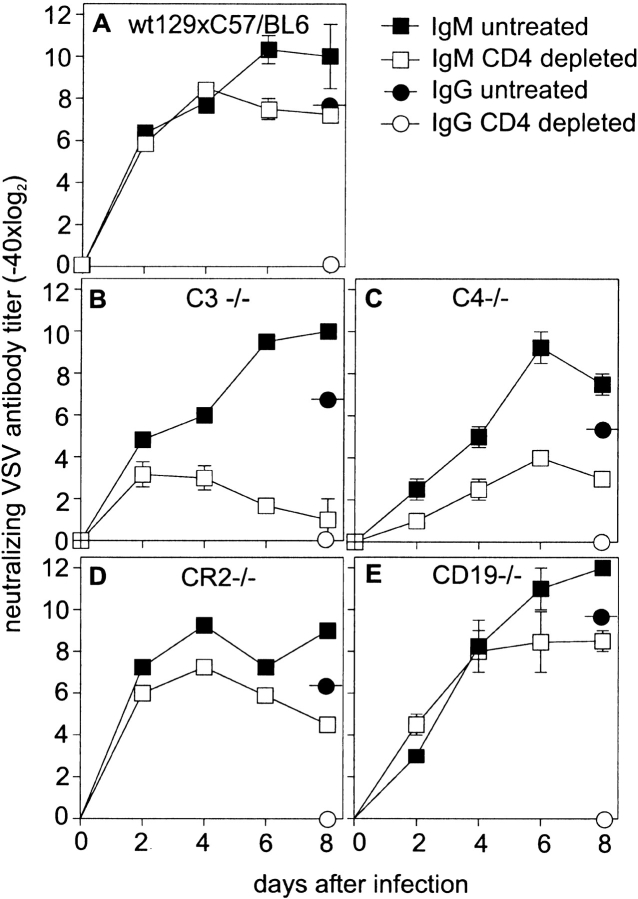
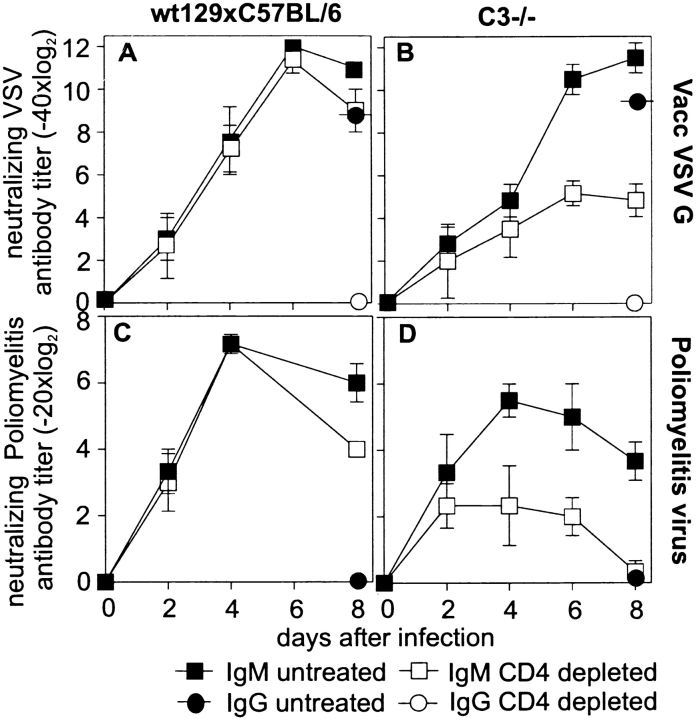
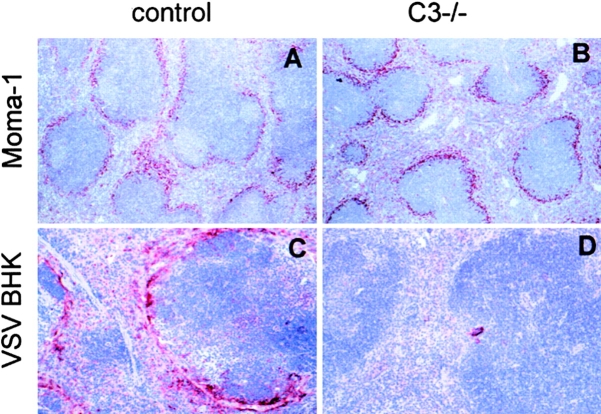
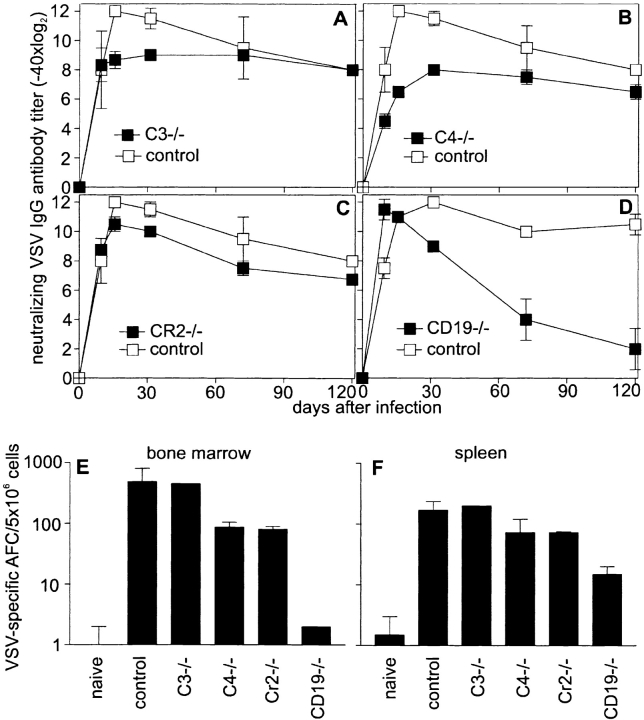
Complement is part of the innate immune system and one of the first lines of host defense against infections. Its importance was evaluated in this study in virus infections in mice deficient either in soluble complement factors (C3(-/-), C4(-/-)) or in the complement signaling complex (complement receptor [CR]2(-/-), CD19(-/-)). The induction of the initial T cell-independent neutralizing immunoglobulin (Ig)M antibody response to vesicular stomatitis virus (VSV), poliomyelitis virus, and recombinant vaccinia virus depended on efficient antigen trapping by CR3 and -4-expressing macrophages of the splenic marginal zone. Neutralizing IgM and IgG antibody responses were largely independent of CR2-mediated stimulation of B cells when mice were infected with live virus. In contrast, immunizations with nonreplicating antigens revealed an important role of B cell stimulation via CR2 in the switch to IgG. The complement cascade was activated after infection with VSV via the classical pathway, and active complement cleavage products augmented the effector function of neutralizing IgM and IgG antibodies to VSV by a factor of 10-100. Absence of the early neutralizing antibody responses, together with the reduced efficiency of neutralizing IgM in C3(-/-) mice, led to a drastically enhanced susceptibility to disease after infection with VSV.
Figures
References
-
- Wessels M.R., Butko P., Ma M., Warren H.B., Lage A.L., Carroll M.C. Studies of group B streptococcal infection in mice deficient in complement component C3 or C4 demonstrate an essential role for complement in both innate and acquired immunity. Proc. Natl. Acad. Sci. USA. 1995;92:11490–11494. - PMC - PubMed
-
- Holland S.M., Gallin J.I. Evaluation of the patient with recurrent bacterial infections. Annu. Rev. Med. 1998;49:185–199. - PubMed
-
- Lachmann P.J., Davies A. Complement and immunity to viruses. Immunol. Rev. 1997;159:69–77. - PubMed
-
- Stoiber H., Clivio A., Dierich M.P. Role of complement in HIV infection. Annu. Rev. Immunol. 1997;15:649–674. - PubMed
-
- Carter R.H., Fearon D.T. CD19lowering the threshold for antigen receptor stimulation of B lymphocytes. Science. 1992;256:105–107. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
