

Eukaryotic translation initiation factor 4AIII (eIF4AIII) is functionally distinct from eIF4AI and eIF4AII
- PMID: 10523622
- PMCID: PMC84727
- DOI: 10.1128/MCB.19.11.7336
Eukaryotic translation initiation factor 4AIII (eIF4AIII) is functionally distinct from eIF4AI and eIF4AII
Abstract
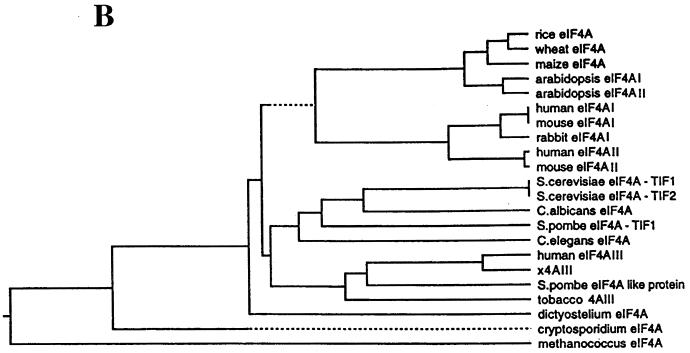
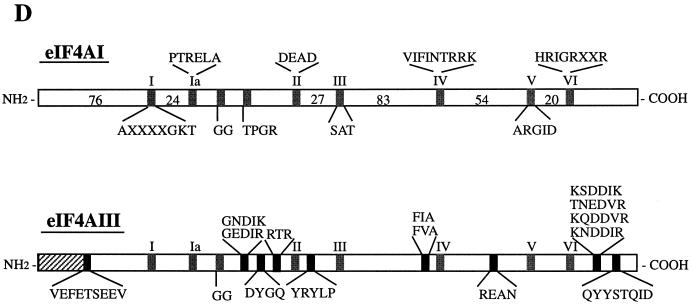
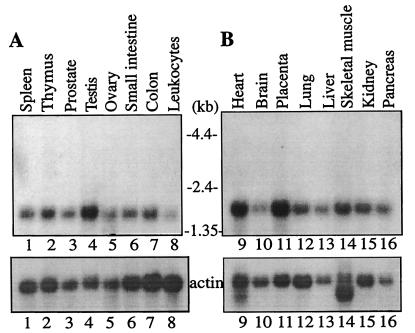
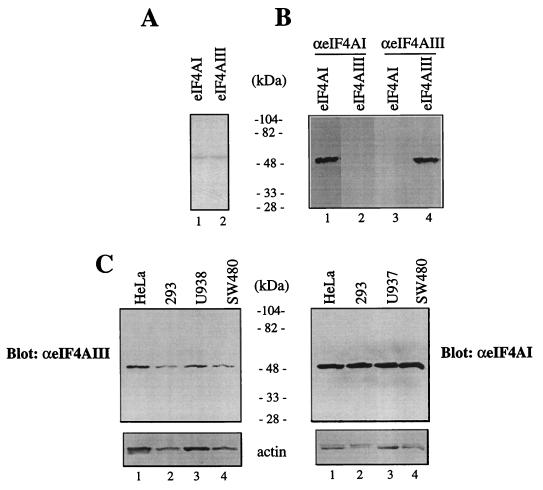
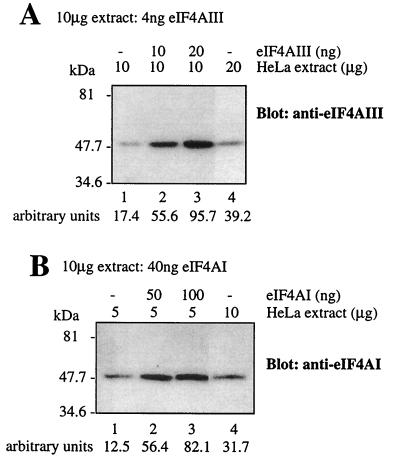
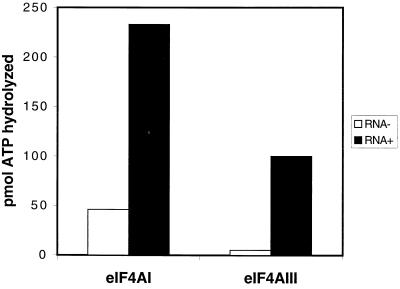
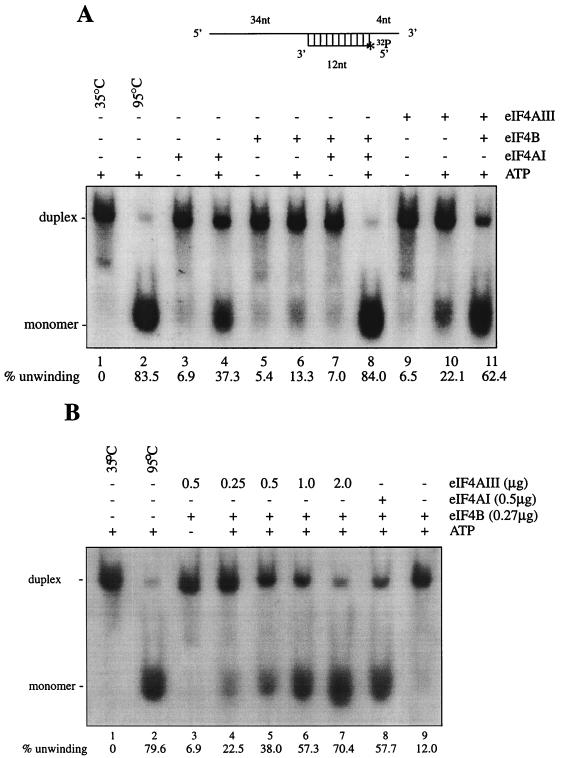
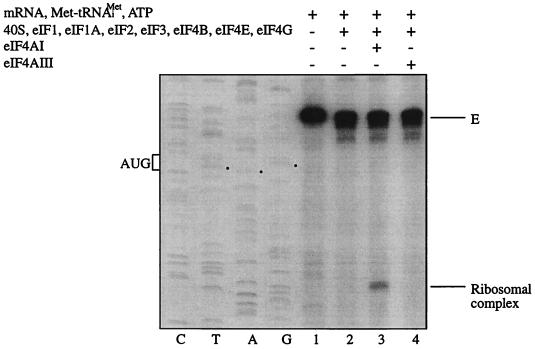
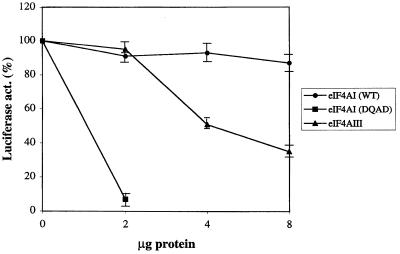
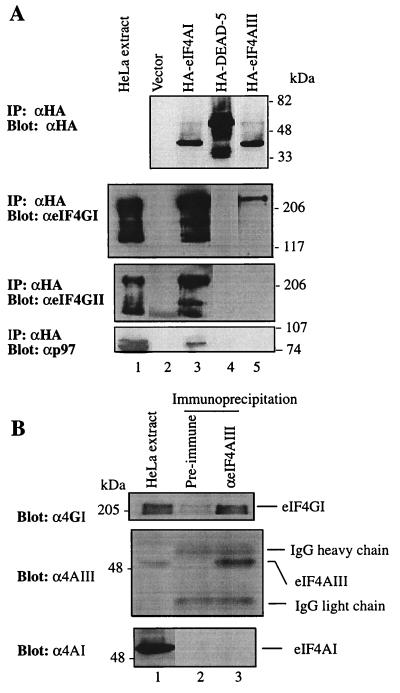
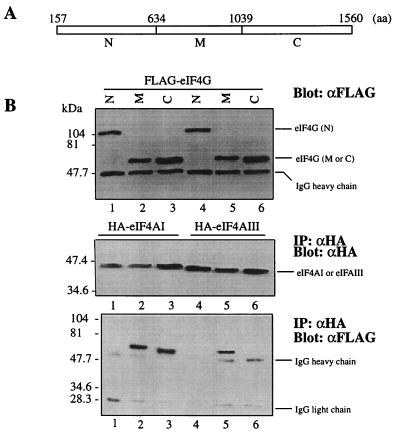
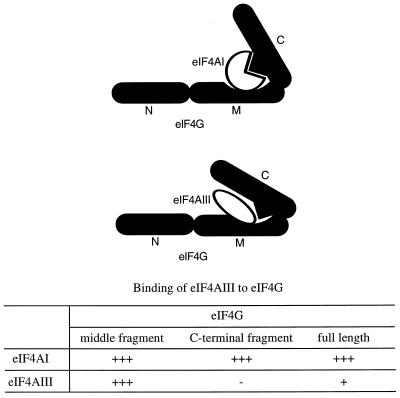
Eukaryotic initiation factor 4A (eIF4A) is an RNA-dependent ATPase and ATP-dependent RNA helicase that is thought to melt the 5' proximal secondary structure of eukaryotic mRNAs to facilitate attachment of the 40S ribosomal subunit. eIF4A functions in a complex termed eIF4F with two other initiation factors (eIF4E and eIF4G). Two isoforms of eIF4A, eIF4AI and eIF4AII, which are encoded by two different genes, are functionally indistinguishable. A third member of the eIF4A family, eIF4AIII, whose human homolog exhibits 65% amino acid identity to human eIF4AI, has also been cloned from Xenopus and tobacco, but its function in translation has not been characterized. In this study, human eIF4AIII was characterized biochemically. While eIF4AIII, like eIF4AI, exhibits RNA-dependent ATPase activity and ATP-dependent RNA helicase activity, it fails to substitute for eIF4AI in an in vitro-reconstituted 40S ribosome binding assay. Instead, eIF4AIII inhibits translation in a reticulocyte lysate system. In addition, whereas eIF4AI binds independently to the middle and carboxy-terminal fragments of eIF4G, eIF4AIII binds to the middle fragment only. These functional differences between eIF4AI and eIF4AIII suggest that eIF4AIII might play an inhibitory role in translation under physiological conditions.
Figures


References
-
- Allen M L, Metz A M, Timmer A T, Rhoads R E, Browning K S. Isolation and sequence of the cDNAs encoding the subunits of the isozyme form of wheat protein synthesis initiation factor 4F. J Biol Chem. 1992;267:23232–23236. - PubMed
-
- Audet R G, Goodchild J, Richter J D. Eukaryotic initiation factor 4A stimulates translation in microinjected Xenopus oocyte. Dev Biol. 1987;121:58–68. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous