

Saccharomyces cerevisiae pol30 (proliferating cell nuclear antigen) mutations impair replication fidelity and mismatch repair
- PMID: 10523669
- PMCID: PMC84846
- DOI: 10.1128/MCB.19.11.7801
Saccharomyces cerevisiae pol30 (proliferating cell nuclear antigen) mutations impair replication fidelity and mismatch repair
Abstract
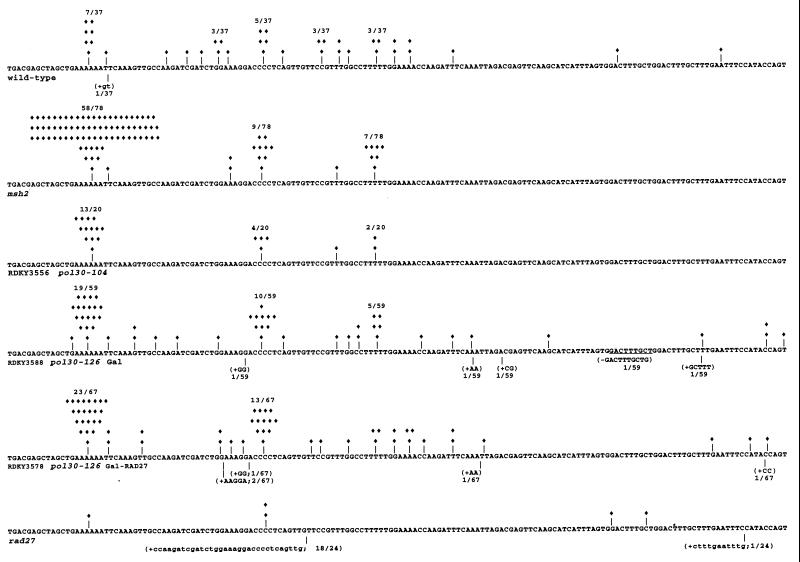
To understand the role of POL30 in mutation suppression, 11 Saccharomyces cerevisiae pol30 mutator mutants were characterized. These mutants were grouped based on their mutagenic defects. Many pol30 mutants harbor multiple mutagenic defects and were placed in more than one group. Group A mutations (pol30-52, -104, -108, and -126) caused defects in mismatch repair (MMR). These mutants exhibited mutation rates and spectra reminiscent of MMR-defective mutants and were defective in an in vivo MMR assay. The mutation rates of group A mutants were enhanced by a msh2 or a msh6 mutation, indicating that MMR deficiency is not the only mutagenic defect present. Group B mutants (pol30-45, -103, -105, -126, and -114) exhibited increased accumulation of either deletions alone or a combination of deletions and duplications (4 to 60 bp). All deletion and duplication breakpoints were flanked by 3 to 7 bp of imperfect direct repeats. Genetic analysis of one representative group B mutant, pol30-126, suggested polymerase slippage as the likely mutagenic mechanism. Group C mutants (pol30-100, -103, -105, -108, and -114) accumulated base substitutions and exhibited synergistic increases in mutation rate when combined with msh6 mutations, suggesting increased DNA polymerase misincorporation as a mutagenic defect. The synthetic lethality between a group A mutant, pol30-104, and rad52 was almost completely suppressed by the inactivation of MSH2. Moreover, pol30-104 caused a hyperrecombination phenotype that was partially suppressed by a msh2 mutation. These results suggest that pol30-104 strains accumulate DNA breaks in a MSH2-dependent manner.
Figures

References
-
- Alani E, Lee S, Kane M F, Griffith J, Kolodner R D. Saccharomyces cerevisiae MSH2, a mispaired base recognition protein, also recognizes Holliday junctions in DNA. J Mol Biol. 1997;265:289–301. - PubMed
-
- Bauer G A, Burgers P M. Protein-protein interactions of yeast DNA polymerase III with mammalian and yeast proliferating cell nuclear antigen (PCNA)/cyclin. Biochim Biophy Acta. 1988;951:274–279. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous