

Analysis of the human thymic perivascular space during aging
- PMID: 10525041
- PMCID: PMC408578
- DOI: 10.1172/JCI7558
Analysis of the human thymic perivascular space during aging
Abstract
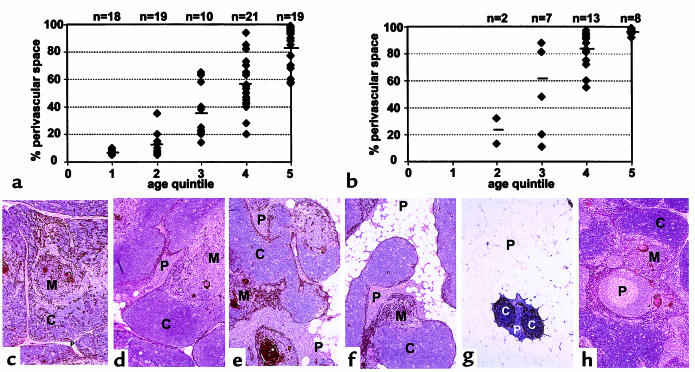
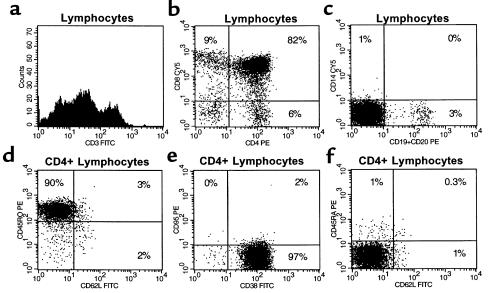
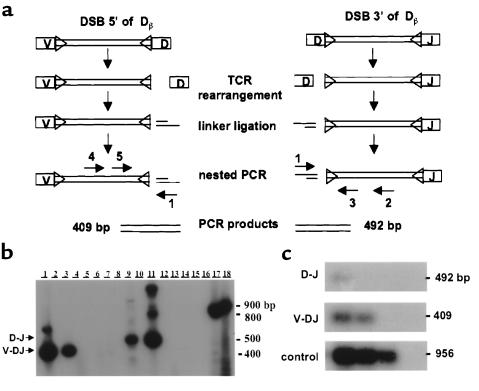
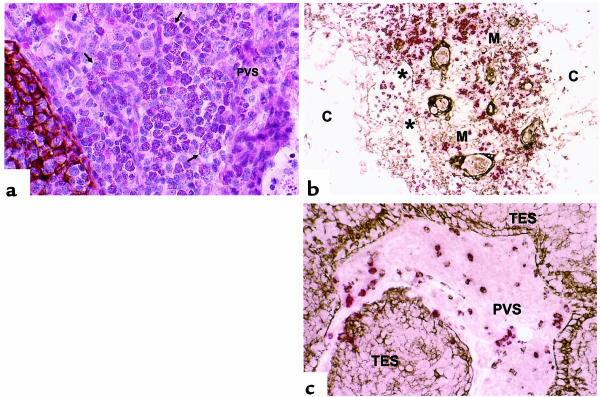
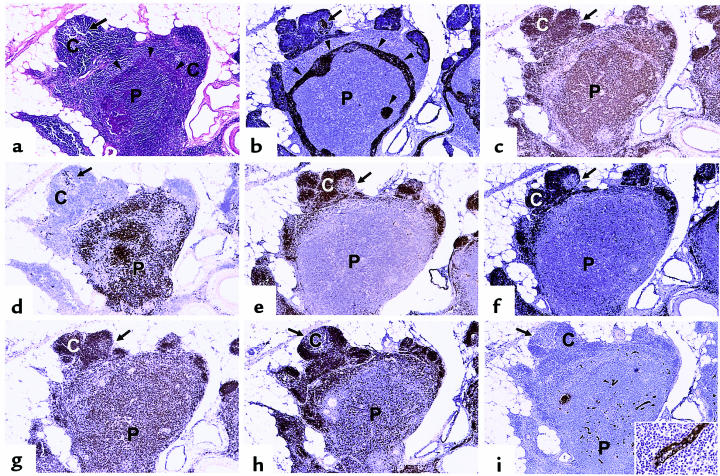
The perivascular space (PVS) of human thymus increases in volume during aging as thymopoiesis declines. Understanding the composition of the PVS is therefore vital to understanding mechanisms of thymic atrophy. We have analyzed 87 normal and 31 myasthenia gravis (MG) thymus tissues from patients ranging in age from newborn to 78 years, using immunohistologic and molecular assays. We confirmed that although thymic epithelial space (TES) volume decreases progressively with age, thymopoiesis with active T-cell receptor gene rearrangement continued normally within the TES into late life. Hematopoietic cells present in the adult PVS include T cells, B cells, and monocytes. Eosinophils are prominent in PVS of infants 2 years of age or younger. In the normal adult and the MG thymus, the PVS includes mature single-positive (CD1a(-) and CD4(+) or CD8(+)) T lymphocytes that express CD45RO, and contains clusters of T cells expressing the TIA-1 cytotoxic granule antigen, suggesting a peripheral origin. PBMCs bind in vitro to MECA-79(+) high endothelial venules present in the PVS, suggesting a mechanism for the recruitment of peripheral cells to thymic PVS. Therefore, in both normal subjects and MG patients, thymic PVS may be a compartment of the peripheral immune system that is not directly involved in thymopoiesis.
Figures
References
-
- Steinmann GG, Klaus B, Muller-Hermelink H-K. The involution of the ageing human thymic epithelium is independent of puberty. A morphometric study. Scand J Immunol. 1985;22:563–575. - PubMed
-
- Steinmann GG. Changes in the human thymus during aging. Curr Top Pathol. 1986;75:43–88. - PubMed
-
- Kato S, Schoefl GI. Microvasculature of normal and involuted mouse thymus: light- and electro-microscopic study. Acta Anat (Basel) 1989;135:1–11. - PubMed
-
- Tamaoki N, Habu S, Kameya T. Thymic lymphoid follicles in autoimmune diseases: histological, histochemical, and electron microscopic studies. Keio J Med. 1971;20:57–68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
