

Differentiation of monocytes to macrophages primes cells for lipopolysaccharide stimulation via accumulation of cytoplasmic nuclear factor kappaB
- PMID: 10531202
- PMCID: PMC96928
- DOI: 10.1128/IAI.67.11.5573-5578.1999
Differentiation of monocytes to macrophages primes cells for lipopolysaccharide stimulation via accumulation of cytoplasmic nuclear factor kappaB
Abstract
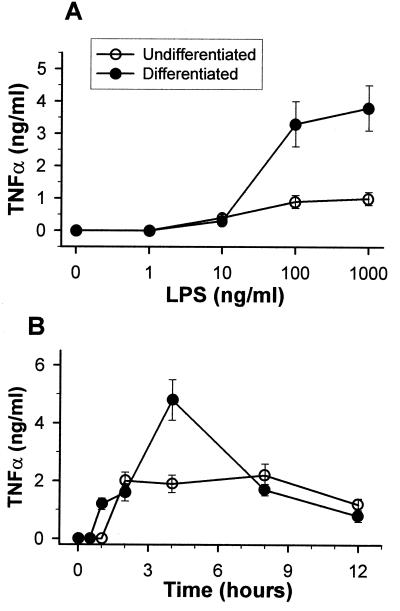
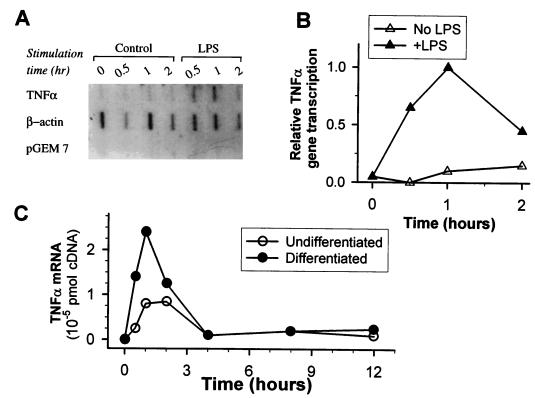
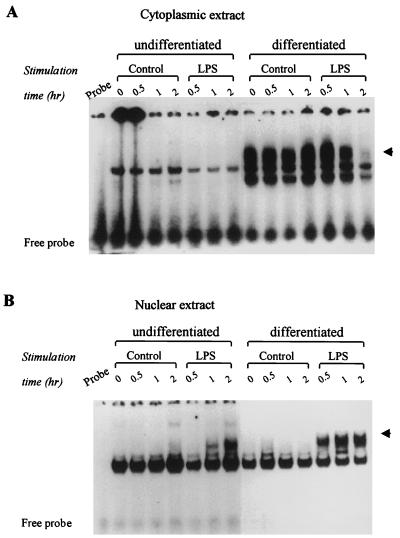
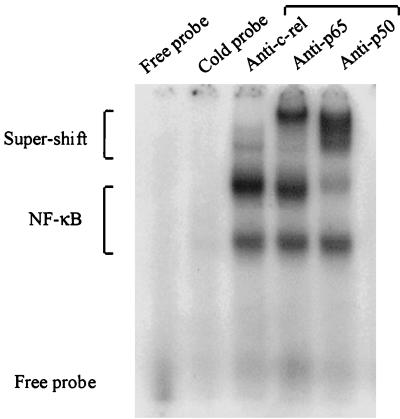
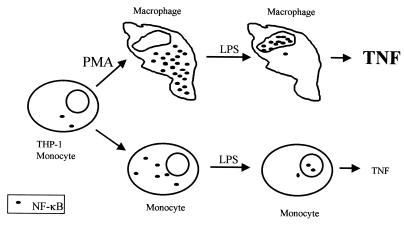
During infection, circulating blood monocytes migrate from the vasculature to the extravascular compartments where they mature into tissue macrophages. The maturation process prepares the cell to actively participate in the inflammatory and the immune responses, and many transcription factors have been found to be involved. Here we report on a novel role for nuclear factor kappaB (NF-kappaB) in this process. Its accumulation in the cytoplasm of differentiated macrophages is responsible for the enhanced ability of the cell to respond to lipopolysaccharide (LPS) stimulation, as determined by tumor necrosis factor alpha (TNF-alpha) secretion. Differentiation of the human monocytic cell line THP-1 into macrophage-like cells was induced by exposure of the cells to phorbol myristate acetate. DNA-bindable NF-kappaB was not detected in the cytoplasm of undifferentiated THP-1 cells but accumulated in the cytoplasm of the cells following differentiation. No TNF-alpha was detected in the media of resting differentiated and nondifferentiated THP-1 cells. Stimulation with LPS of differentiated cells induced the production of higher levels of TNF-alpha than stimulation of nondifferentiated cells. This hyperresponsiveness to LPS was found in the mRNA and secreted TNF-alpha levels. Furthermore, stimulation with LPS induced the translocation of NF-kappaB from the cytoplasm into the nucleus. This translocation process was more rapid in the differentiated cells than in the nondifferentiated cells, and the resultant accumulated levels of NF-kappaB in the nucleus were higher. The DNA-bindable NF-kappaB was identified as a heterodimer of p65 and p50. The results suggest that NF-kappaB accumulation in the cytoplasm during maturation of monocytes to macrophages primes the cells for enhanced responsiveness to LPS and results in the rapid secretion of inflammatory mediators, such as TNF-alpha, by mature macrophages following LPS challenge.
Figures
References
-
- Abmayer S M, Workman J L. Preparation of nuclear and cytoplasmic extracts from mammalian cells. In: Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: Greene Publishing and Wiley Interscience; 1990. pp. 12.1.1–12.1.9.
-
- Auger M J, Ross J A. The biology of the macrophage. In: Lewis C E, McGee J O, editors. The macrophage. New York, N.Y: IRL Press; 1992. pp. 3–74.
-
- Baeuerle P A. The inducible transcription activator NF-κB: regulation by distinct protein subunits. Biochim Biophys Acta. 1991;1072:63–80. - PubMed
-
- Bondeson J, Browne K A, Brennan F M, Foxwell B M J, Feldmann M. Selective regulation of cytokine induction by adenoviral gene transfer of IκB into human macrophages: lipopolysaccharide-induced, but not zymosan-induced, proinflammatory cytokines are inhibited, but IL-10 is nuclear factor-κB independent. J Immunol. 1999;162:2939–2945. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
