

Lysine residue 117 of the FasG adhesin of enterotoxigenic Escherichia coli is essential for binding of 987P fimbriae to sulfatide
- PMID: 10531225
- PMCID: PMC96951
- DOI: 10.1128/IAI.67.11.5755-5761.1999
Lysine residue 117 of the FasG adhesin of enterotoxigenic Escherichia coli is essential for binding of 987P fimbriae to sulfatide
Abstract
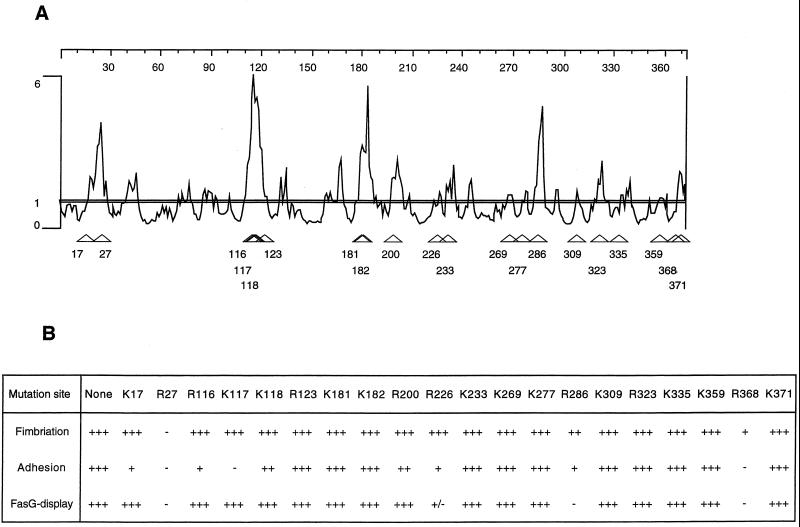
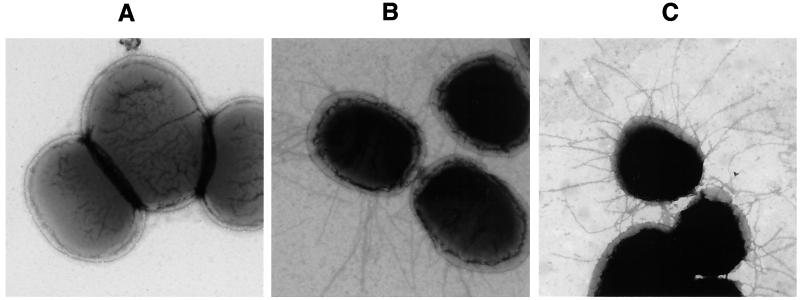
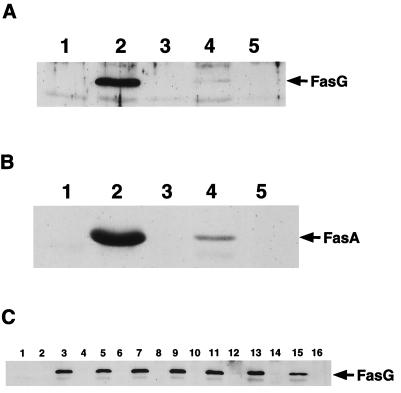
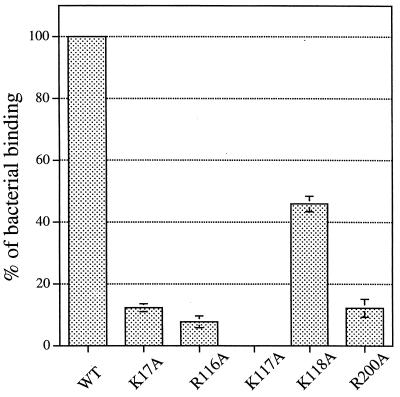
The FasG subunit of the 987P fimbriae of enterotoxigenic strains of Escherichia coli was previously shown to mediate fimbrial binding to a glycoprotein and a sulfatide receptor on intestinal brush borders of piglets. Moreover, the 987P adhesin FasG is required for fimbrial expression, since fasG null mutants are nonfimbriated. In this study, fasG was modified by site-directed mutagenesis to study its sulfatide binding properties. Twenty single mutants were generated by replacing positively charged lysine (K) or arginine (R) residues with small, nonpolar alanine (A) residues. Reduced levels of binding to sulfatide-containing liposomes correlated with reduced fimbriation and FasG surface display in four fasG mutants (R27A, R286A, R226A, and R368). Among the 16 remaining normally fimbriated mutants with wild-type levels of surface-exposed FasG, only one mutant (K117A) did not interact at all with sulfatide-containing liposomes. Four mutants (K117A, R116A, K118A, and R200A) demonstrated reduced binding to such liposomes. Since complete phenotypic dissociation between the structure and specific function of 987P was observed only with mutant K117A, this residue is proposed to play an essential role in the FasG-sulfatide interaction, possibly communicating with the sulfate group of sulfatide by hydrogen bonding and/or salt bridge formation. Residues K17, R116, K118, and R200 may stabilize this interaction.
Figures
References
-
- Choi, B. K., and D. M. Schifferli. 1999. Unpublished data.
-
- Cunningham B C, Wells J A. High-resolution epitope mapping of hGH-receptor interactions by alanine-scanning mutagenesis. Science. 1989;244:1081–1085. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
