

Identification of regions of the chromosome of Neisseria meningitidis and Neisseria gonorrhoeae which are specific to the pathogenic Neisseria species
- PMID: 10531275
- PMCID: PMC97001
- DOI: 10.1128/IAI.67.11.6119-6129.1999
Identification of regions of the chromosome of Neisseria meningitidis and Neisseria gonorrhoeae which are specific to the pathogenic Neisseria species
Abstract
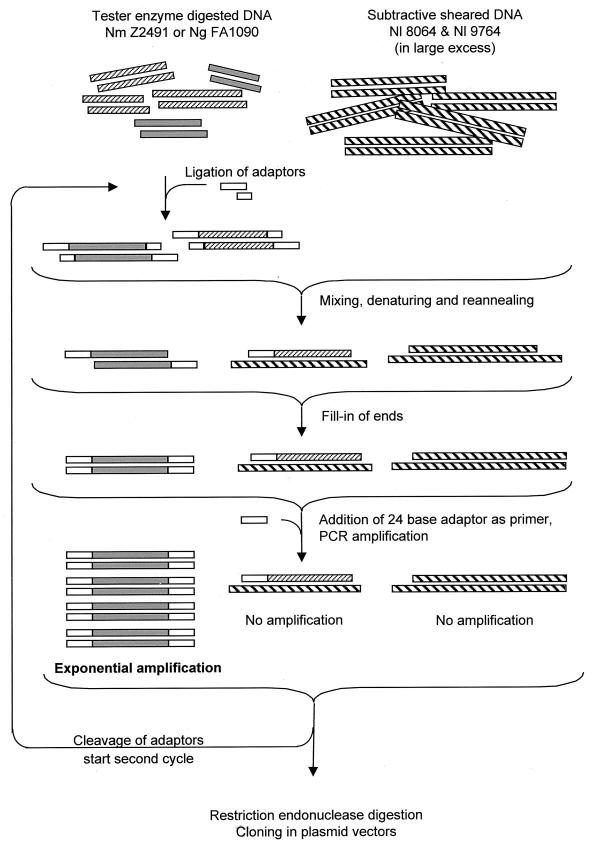
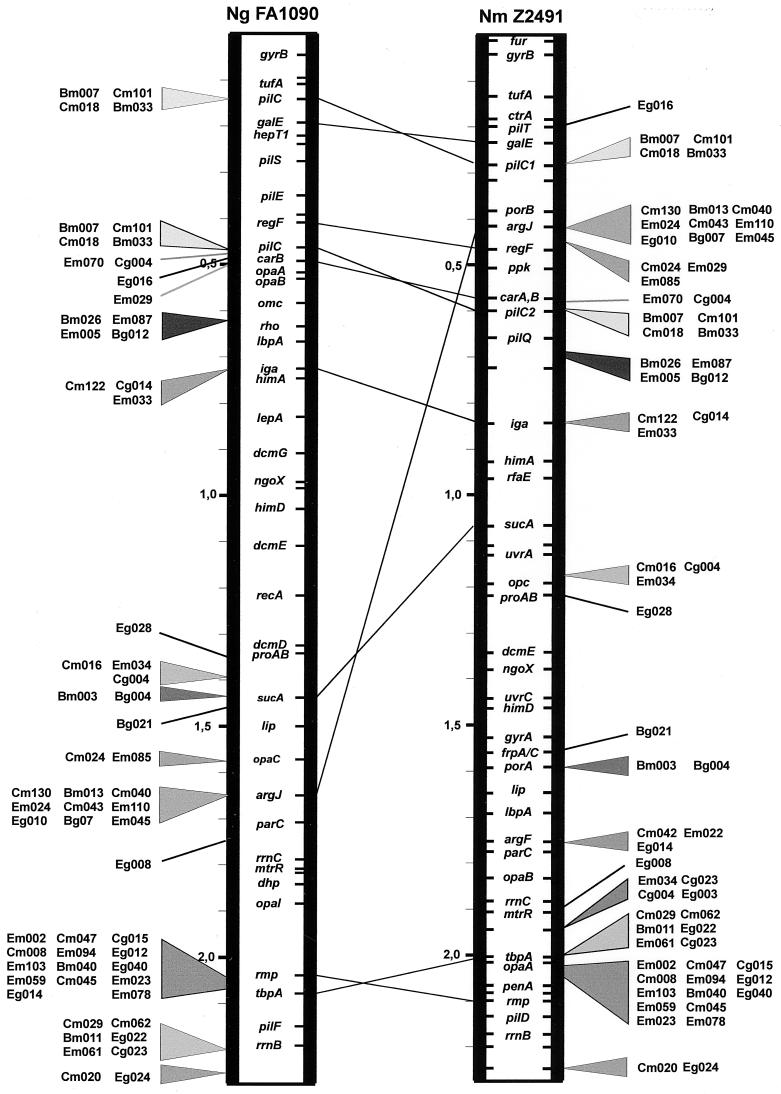
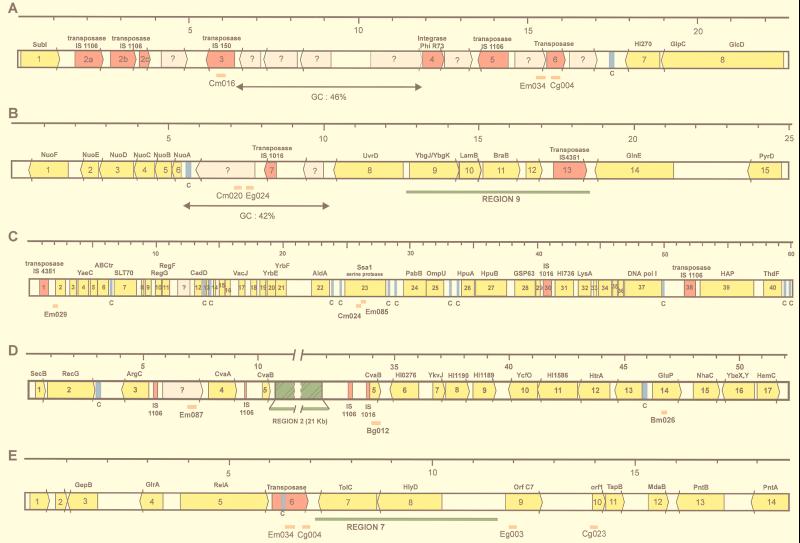
Neisseria meningitidis and Neisseria gonorrhoeae give rise to dramatically different diseases. Their interactions with the host, however, do share common characteristics: they are both human pathogens which do not survive in the environment and which colonize and invade mucosa at their port of entry. It is therefore likely that they have common properties that might not be found in nonpathogenic bacteria belonging to the same genetically related group, such as Neisseria lactamica. Their common properties may be determined by chromosomal regions found only in the pathogenic Neisseria species. To address this issue, we used a previously described technique (C. R. Tinsley and X. Nassif, Proc. Natl. Acad. Sci. USA 93:11109-11114, 1996) to identify sequences of DNA specific for pathogenic neisseriae and not found in N. lactamica. Sequences present in N. lactamica were physically subtracted from the N. meningitidis Z2491 sequence and also from the N. gonorrhoeae FA1090 sequence. The clones obtained from each subtraction were tested by Southern blotting for their reactivity with the three species, and only those which reacted with both N. meningitidis and N. gonorrhoeae (i.e., not specific to either one of the pathogens) were further investigated. In a first step, these clones were mapped onto the chromosomes of both N. meningitidis and N. gonorrhoeae. The majority of the clones were arranged in clusters extending up to 10 kb, suggesting the presence of chromosomal regions common to N. meningitidis and N. gonorrhoeae which distinguish these pathogens from the commensal N. lactamica. The sequences surrounding these clones were determined from the N. meningitidis genome-sequencing project. Several clones corresponded to previously described factors required for colonization and survival at the port of entry, such as immunoglobulin A protease and PilC. Others were homologous to virulence-associated proteins in other bacteria, demonstrating that the subtractive clones are capable of pinpointing chromosomal regions shared by N. meningitidis and N. gonorrhoeae which are involved in common aspects of the host interaction of both pathogens.
Figures

Similar articles
-
Genome analysis and strain comparison of correia repeats and correia repeat-enclosed elements in pathogenic Neisseria.J Bacteriol. 2002 Nov;184(22):6163-73. doi: 10.1128/JB.184.22.6163-6173.2002. J Bacteriol. 2002. PMID: 12399486 Free PMC article.
-
Molecular and biological analysis of eight genetic islands that distinguish Neisseria meningitidis from the closely related pathogen Neisseria gonorrhoeae.Infect Immun. 2000 Apr;68(4):2082-95. doi: 10.1128/IAI.68.4.2082-2095.2000. Infect Immun. 2000. PMID: 10722605 Free PMC article.
-
Identification and characterization of specific sequences encoding pathogenicity associated proteins in the genome of commensal Neisseria species.FEMS Microbiol Lett. 1995 Jan 15;125(2-3):255-63. doi: 10.1111/j.1574-6968.1995.tb07366.x. FEMS Microbiol Lett. 1995. PMID: 7875573
-
Population genomics: diversity and virulence in the Neisseria.Curr Opin Microbiol. 2008 Oct;11(5):467-71. doi: 10.1016/j.mib.2008.09.002. Epub 2008 Oct 14. Curr Opin Microbiol. 2008. PMID: 18822386 Free PMC article. Review.
-
Do pathogenic neisseriae need several ways to modify the host cell cytoskeleton?Microbes Infect. 2000 Jun;2(7):821-7. doi: 10.1016/s1286-4579(00)90367-8. Microbes Infect. 2000. PMID: 10955963 Review.
Cited by
-
Genome analysis and strain comparison of correia repeats and correia repeat-enclosed elements in pathogenic Neisseria.J Bacteriol. 2002 Nov;184(22):6163-73. doi: 10.1128/JB.184.22.6163-6173.2002. J Bacteriol. 2002. PMID: 12399486 Free PMC article.
-
A putatively phase variable gene (dca) required for natural competence in Neisseria gonorrhoeae but not Neisseria meningitidis is located within the division cell wall (dcw) gene cluster.J Bacteriol. 2001 Feb;183(4):1233-41. doi: 10.1128/JB.183.4.1233-1241.2001. J Bacteriol. 2001. PMID: 11157935 Free PMC article.
-
Identification, characterization, and variable expression of a naturally occurring inhibitor protein of IS1106 transposase in clinical isolates of Neisseria meningitidis.Infect Immun. 2001 Dec;69(12):7425-36. doi: 10.1128/IAI.69.12.7425-7436.2001. Infect Immun. 2001. PMID: 11705917 Free PMC article.
-
A new confirmatory Neisseria gonorrhoeae real-time PCR assay targeting the porA pseudogene.Eur J Clin Microbiol Infect Dis. 2004 Sep;23(9):705-10. doi: 10.1007/s10096-004-1170-0. Epub 2004 Jul 10. Eur J Clin Microbiol Infect Dis. 2004. PMID: 15248092
-
Representational difference analysis between Afa/Dr diffusely adhering Escherichia coli and nonpathogenic E. coli K-12.Infect Immun. 2002 Oct;70(10):5503-11. doi: 10.1128/IAI.70.10.5503-5511.2002. Infect Immun. 2002. PMID: 12228276 Free PMC article.
References
-
- Aiba H, Baba T, Fujita K, Hayashi K, Inada T, Isono K, Itoh T, Kasai H, Kashimoto K, Kimura S, Kitakawa M, Kitagawa M, Makino K, Miki T, Mizobuchi K, Mori H, Mori T, Motomura K, Nakade S, Nakamura Y, Nashimoto H, Nishio Y, Oshima T, Saito N, Sampei G, Seki Y, Sivasundaram S, Tagami H, Takeda J, Takemoto K, Takeuchi Y, Wada C, Yamamoto Y, Horiuchi T. A 570-kb DNA sequence of the Escherichia coli K-12 genome corresponding to the 28.0-40.1 min region on the linkage map. DNA Res. 1996;3:363–377. - PubMed
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Current protocols in molecular biology. Vol. 1. New York, N.Y: Greene Publishing Associates and Wiley-Interscience; 1989.
-
- Baida G E, Kuzmin N P. Cloning and primary structure of a new hemolysin gene from Bacillus cereus. Biochim Biophys Acta. 1995;1264:151–154. - PubMed
-
- Barlow A K, Heckels J E, Clarke I N. The class 1 outer membrane protein of Neisseria meningitidis: gene sequence and structural and immunological similarities to gonococcal porins. Mol Microbiol. 1989;3:131–139. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
