

Involvement of an active efflux system in the natural resistance of Pseudomonas aeruginosa to aminoglycosides
- PMID: 10543738
- PMCID: PMC89534
- DOI: 10.1128/AAC.43.11.2624
Involvement of an active efflux system in the natural resistance of Pseudomonas aeruginosa to aminoglycosides
Abstract
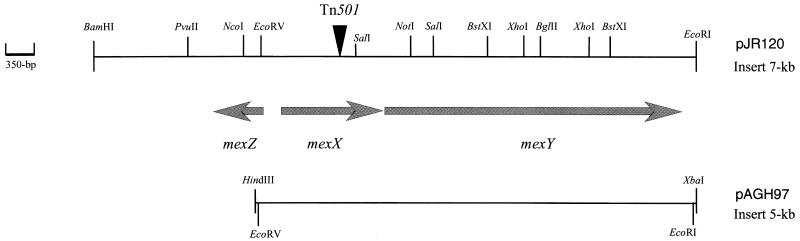
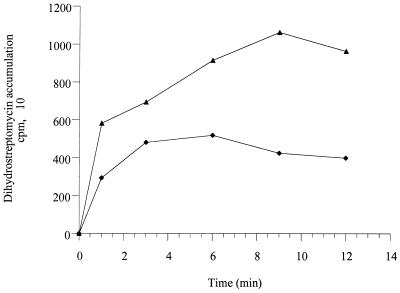
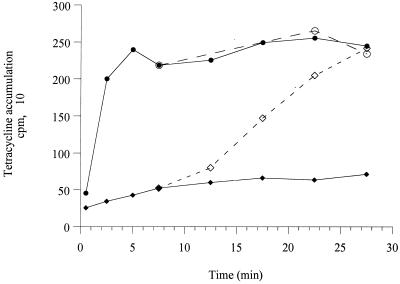
A mutant, named 11B, hypersusceptible to aminoglycosides, tetracycline, and erythromycin was isolated after Tn501 insertion mutagenesis of Pseudomonas aeruginosa PAO1. Cloning and sequencing experiments showed that 11B was deficient in an, at that time, unknown active efflux system that contains homologs of MexAB. This locus also contained a putative regulatory gene, mexZ, transcribed divergently from the efflux operon. Introduction of a recombinant plasmid that carries the genes of the efflux system restored the resistance of 11B to parental levels, whereas overexpression of these genes strongly increased the MICs of substrate antibiotics for the PAO1 host. Antibiotic accumulation studies confirmed that this new system is an energy-dependent active efflux system that pumps out aminoglycosides. Furthermore, this system appeared to function with an outer membrane protein, OprM. While the present paper was being written and reviewed, genes with a sequence identical to our pump genes, mexXY of P. aeruginosa, have been reported to increase resistance to erythromycin, fluoroquinolones, and organic cations in Escherichia coli hosts, although efflux of aminoglycosides was not examined (Mine et al., Antimicrob. Agents Chemother. 43:415-417, 1999). Our study thus shows that the MexXY system plays an important role in the intrinsic resistance of P. aeruginosa to aminoglycosides. Although overexpression of MexXY increased the level of resistance to fluoroquinolones, disruption of the mexXY operon in P. aeruginosa had no detectable effect on susceptibility to these agents.
Figures
References
-
- Balows A, Hausler W J Jr, Herrmann K L, Isenberg H D, Shadomy H J, editors. Manual of clinical microbiology. 5th ed. Washington, D.C.: ASM Press; 1991.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
