

Inhibition of interleukin 2 signaling and signal transducer and activator of transcription (STAT)5 activation during T cell receptor-mediated feedback inhibition of T cell expansion
- PMID: 10544198
- PMCID: PMC2195684
- DOI: 10.1084/jem.190.9.1263
Inhibition of interleukin 2 signaling and signal transducer and activator of transcription (STAT)5 activation during T cell receptor-mediated feedback inhibition of T cell expansion
Abstract
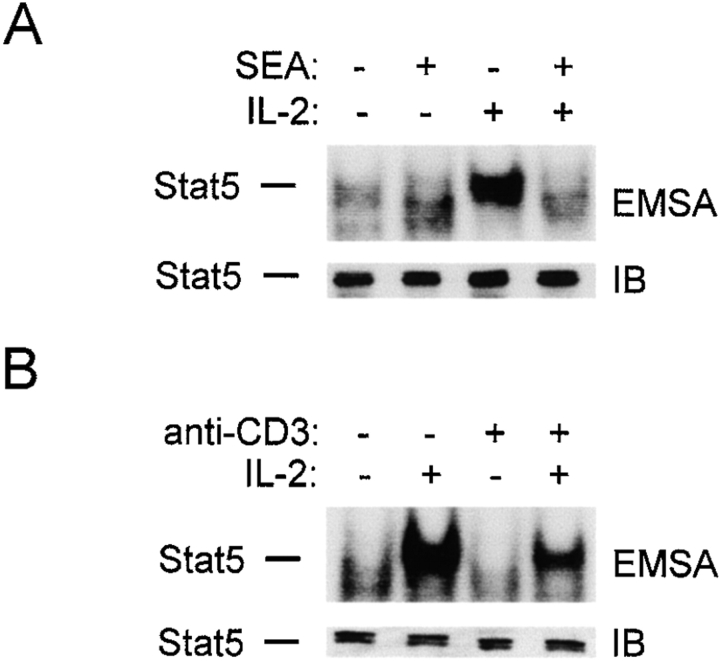
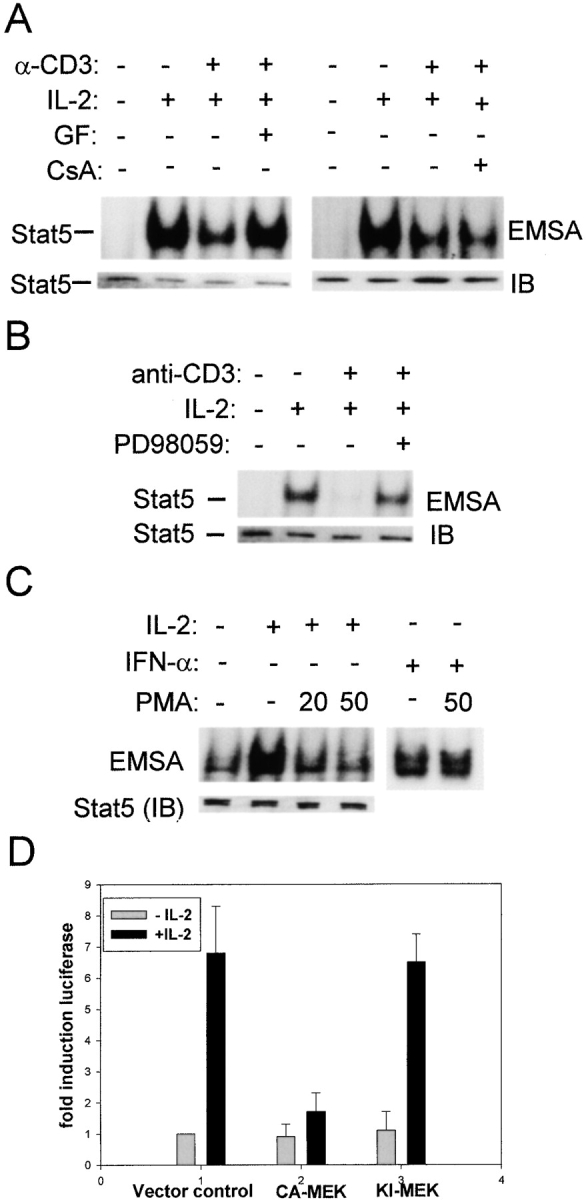
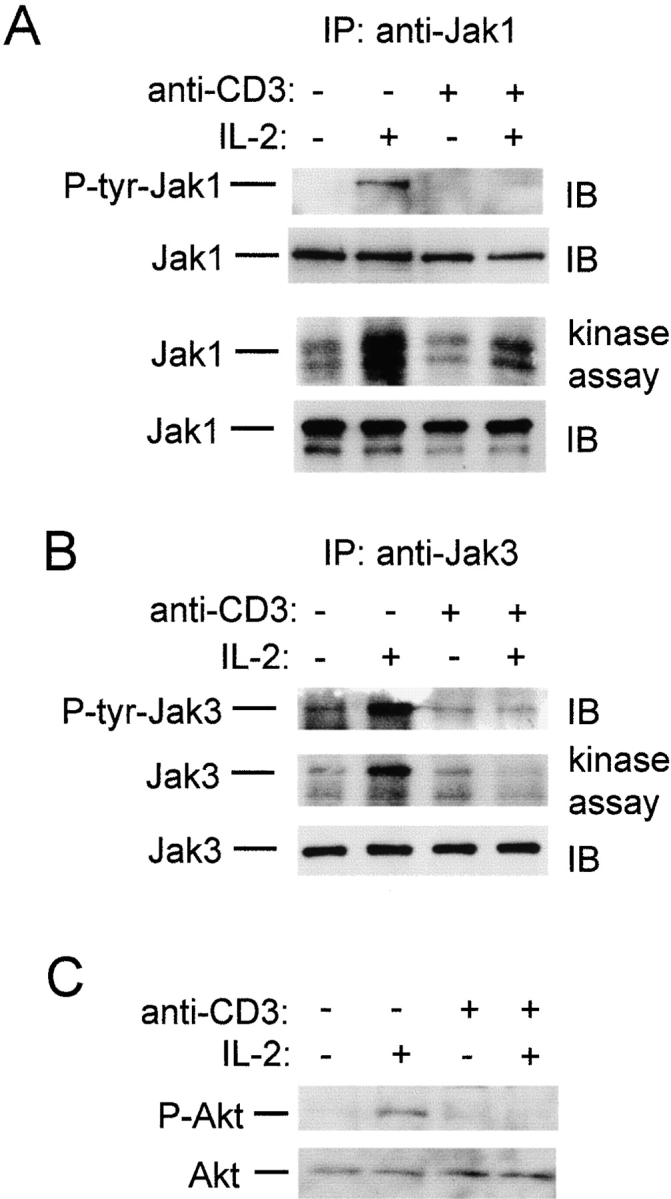
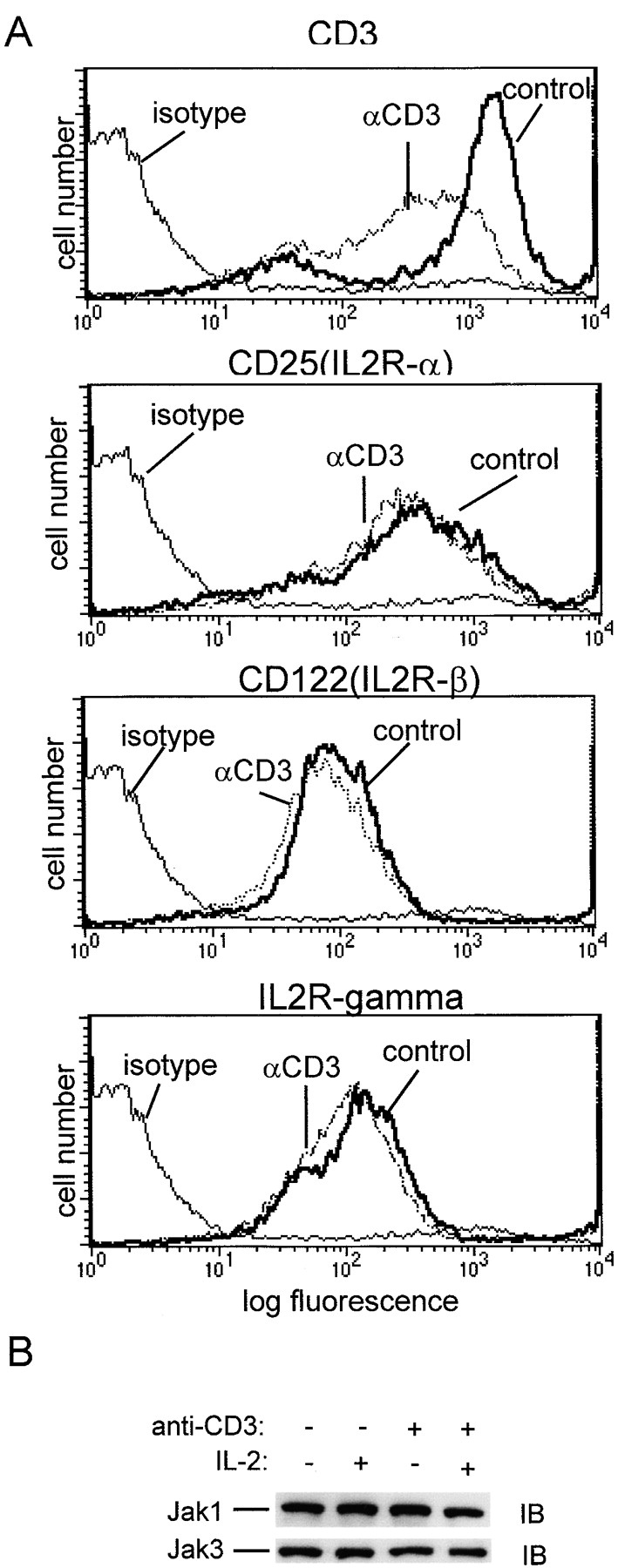
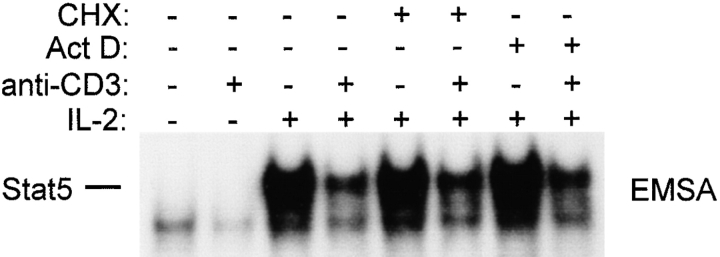
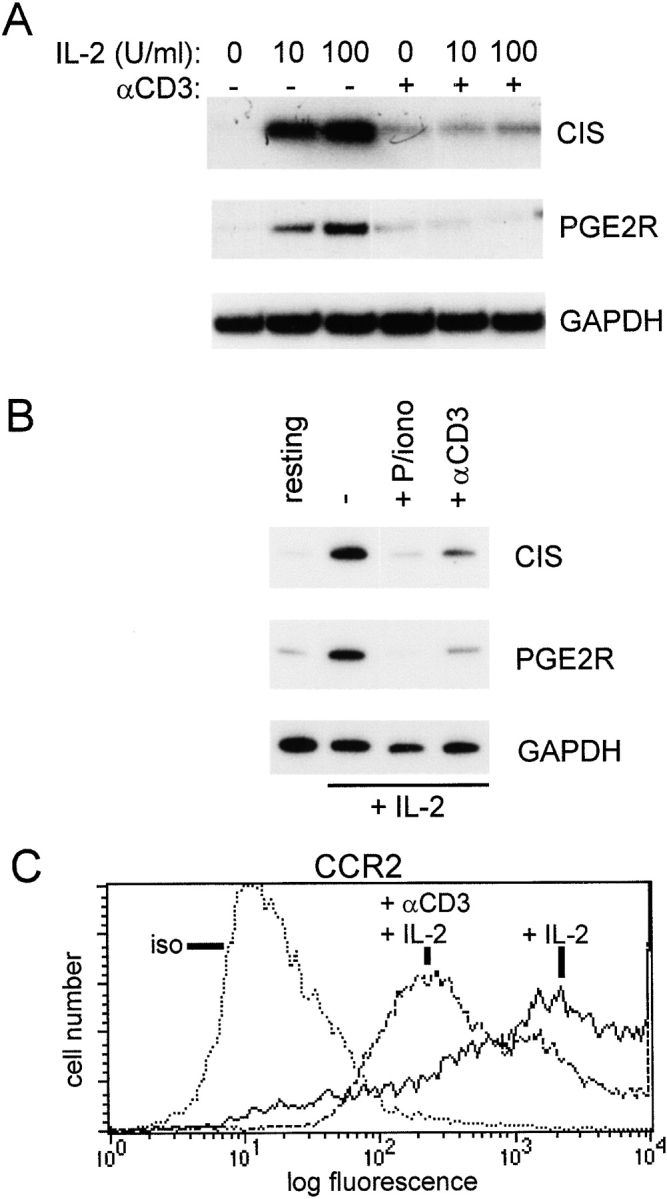
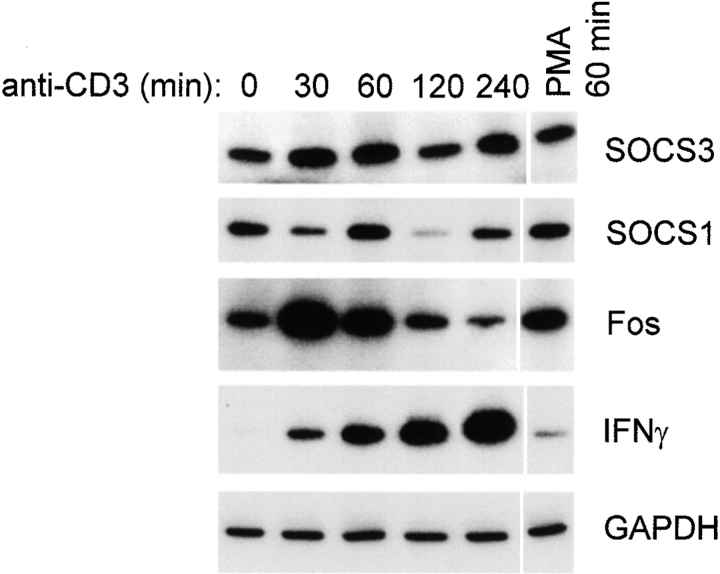
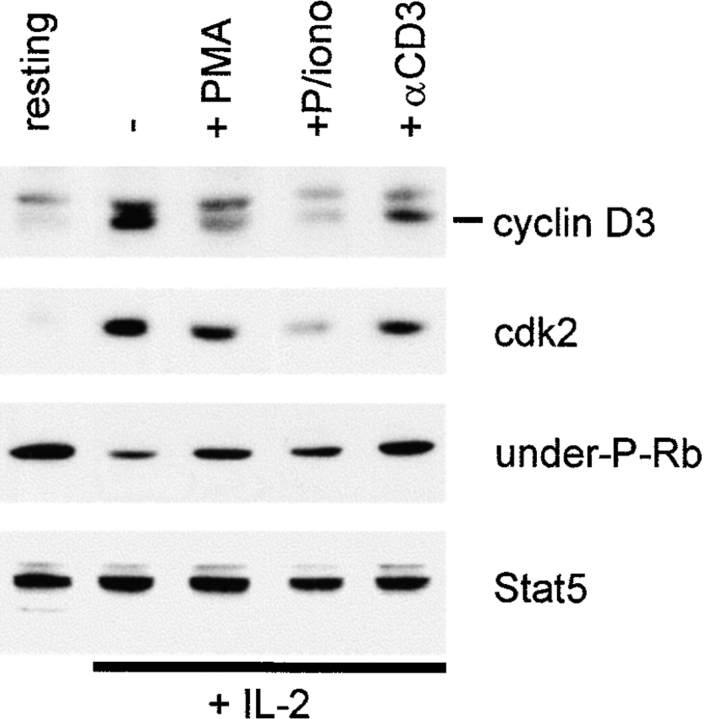
Limitation of clonal expansion of activated T cells is necessary for immune homeostasis, and is achieved by growth arrest and apoptosis. Growth arrest and apoptosis can occur passively secondary to cytokine withdrawal, or can be actively induced by religation of the T cell receptor (TCR) in previously activated proliferating T cells. TCR-induced apoptosis appears to require prior growth arrest, and is mediated by death receptors such as Fas. We tested whether TCR religation affects T cell responses to interleukin (IL)-2, a major T cell growth and survival factor. TCR ligation in activated primary human T cells blocked IL-2 induction of signal transducer and activator of transcription (STAT)5 DNA binding, phosphorylation of STAT5, Janus kinase (Jak)1, Jak3, and Akt, and kinase activity of Jak1 and Jak3. Inhibition was mediated by the mitogen-activated protein kinase kinase (MEK)-extracellular stimulus-regulated kinase (ERK) signaling pathway, similar to the mechanism of inhibition of IL-6 signaling we have described previously. TCR ligation blocked IL-2 activation of genes and cell cycle regulatory proteins, and suppressed cell proliferation and expansion. These results identify TCR-induced inhibition of IL-2 signaling as a novel mechanism that underlies antigen-mediated feedback limitation of T cell expansion, and suggest that modulation of cytokine activity by antigen receptor signals plays an important role in the regulation of lymphocyte function.
Figures

References
-
- Kotzin B.L., Leung D.Y., Kappler J., Marrack P. Superantigens and their potential role in human disease. Adv. Immunol. 1993;54:99–166. - PubMed
-
- Oxenius A., Zinkernagel R.M., Hengartner H. Comparison of activation versus induction of unresponsiveness of virus-specific CD4+ and CD8+ T cells upon acute versus persistent viral infection. Immunity. 1998;9:449–457. - PubMed
-
- Van Parijs L., Abbas A.K. Homeostasis and self-tolerance in the immune systemturning lymphocytes off. Science. 1998;280:243–248. - PubMed
-
- Kuroda K., Yagi J., Imanishi K., Yan X.J., Li X.Y., Fujimaki W., Kato H., Miyoshi-Akiyama T., Kumazawa Y., Abe H., Uchiyama T. Implantation of IL-2-containing osmotic pump prolongs the survival of superantigen-reactive T cells expanded in mice injected with bacterial superantigen. J. Immunol. 1996;157:1422–1431. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
