

The chemokine SDF-1 stimulates integrin-mediated arrest of CD34(+) cells on vascular endothelium under shear flow
- PMID: 10545519
- PMCID: PMC409822
- DOI: 10.1172/JCI7615
The chemokine SDF-1 stimulates integrin-mediated arrest of CD34(+) cells on vascular endothelium under shear flow
Abstract
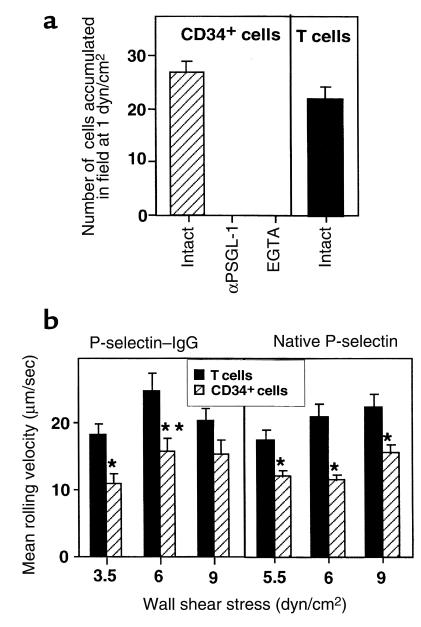
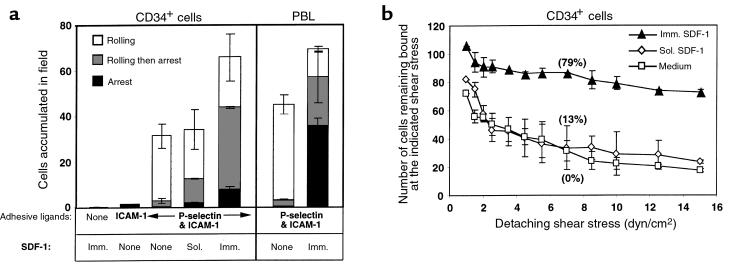
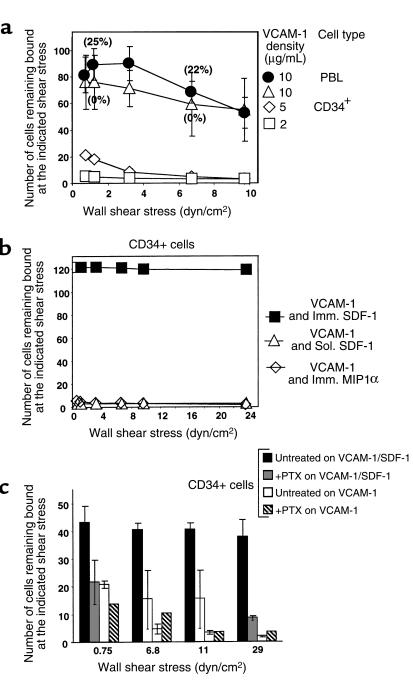
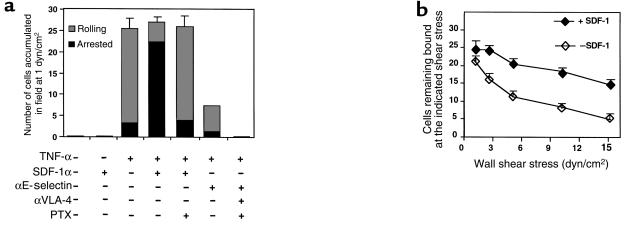
The chemokine SDF-1 plays a central role in the repopulation of the bone marrow (BM) by circulating CD34(+) progenitors, but the mechanisms of its action remain obscure. To extravasate to target tissue, a blood-borne cell must arrest firmly on vascular endothelium. Murine hematopoietic progenitors were recently shown in vivo to roll along BM microvessels that display selectins and integrins. We now show that SDF-1 is constitutively expressed by human BM endothelium. In vitro, human CD34(+) cells establish efficient rolling on P-selectin, E-selectin, and the CD44 ligand hyaluronic acid under physiological shear flow. ICAM-1 alone did not tether CD34(+) cells under flow, but, in the presence of surface-bound SDF-1, CD34(+) progenitors rolling on endothelial selectin rapidly developed firm adhesion to the endothelial surface, mediated by an interaction between ICAM-1 and its integrin ligand, which coimmobilized with SDF-1. Human CD34(+) cells accumulated efficiently on TNF-activated human umbilical cord endothelial cells in the absence of SDF-1, but they required immobilized SDF-1 to develop firm integrin-mediated adhesion and spreading. In the absence of selectins, SDF-1 also promoted VLA-4-mediated, Gi protein-dependent tethering and firm adhesion to VCAM-1 under shear flow. To our knowledge, this is the first demonstration that SDF-1 expressed on vascular endothelium is crucial for translating rolling adhesion of CD34(+) progenitors into firm adhesion by increasing the adhesiveness of the integrins VLA-4 and LFA-1 to their respective endothelial ligands, VCAM-1 and ICAM-1.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
