

Defective mutations in the insulin promoter factor-1 (IPF-1) gene in late-onset type 2 diabetes mellitus
- PMID: 10545531
- PMCID: PMC409821
- DOI: 10.1172/JCI7469
Defective mutations in the insulin promoter factor-1 (IPF-1) gene in late-onset type 2 diabetes mellitus
Abstract
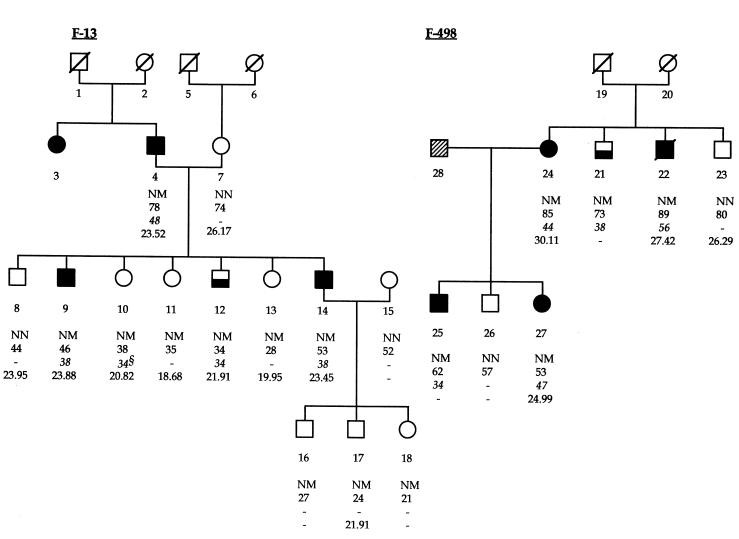
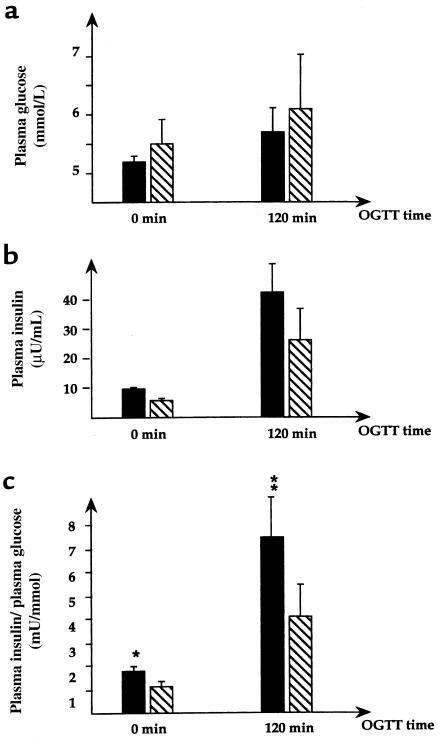
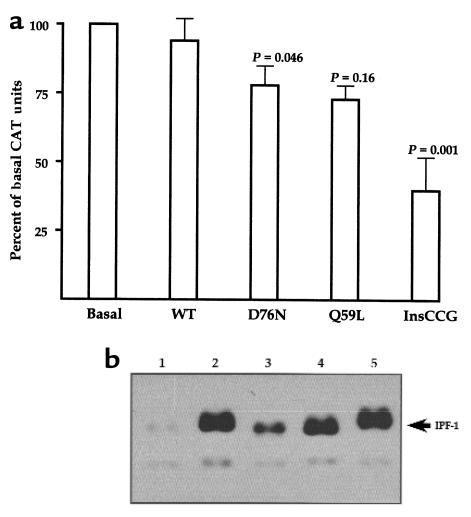
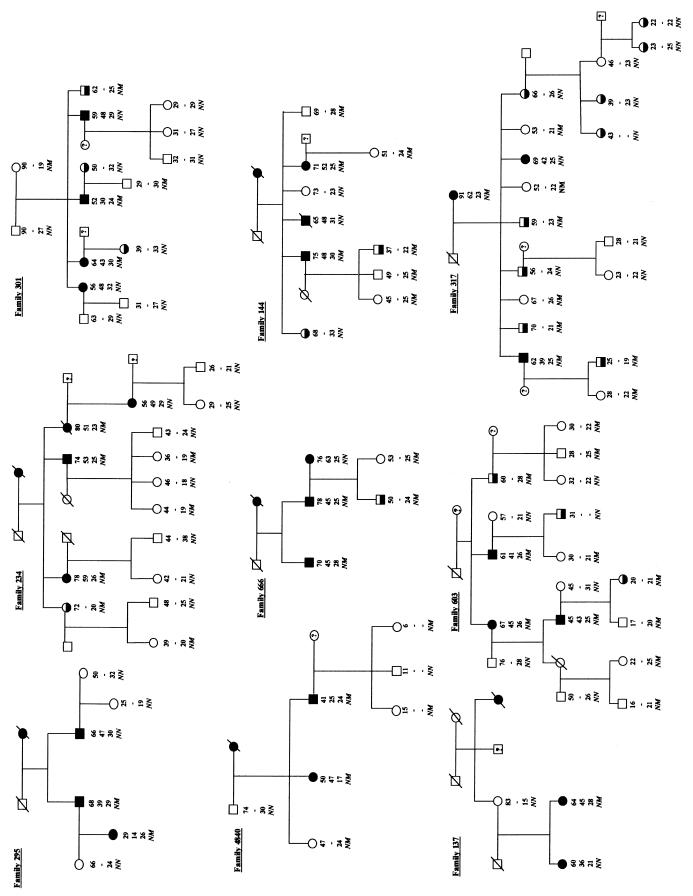
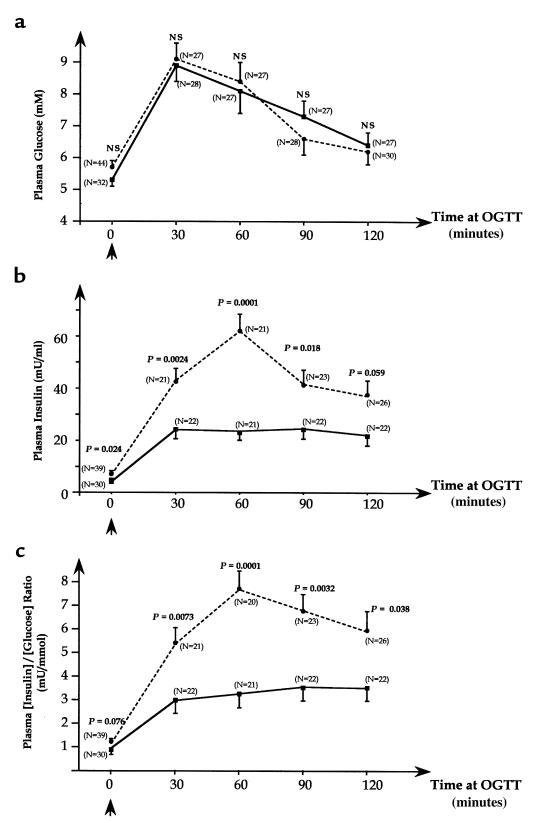
Type 2 diabetes mellitus is a common disabling disease with onset in middle-aged individuals, caused by an imbalance between insulin production and action. Genetic studies point to major genetic components, but, with the exception of maturity-onset diabetes of the young (MODY), specific diabetes susceptibility genes remain to be identified. Recent studies showed that a dominant negative mutation in the insulin promoter factor-1 (IPF-1), a pancreatic beta-cell specific transcription factor, causes pancreatic agenesis and MODY. Thus, we investigated 192 French, non-MODY type 2 diabetic families for mutations in IPF-1. We identified 3 novel IPF-1 mutations, including 2 substitutions (Q59L and D76N) and an in-frame proline insertion (InsCCG243). Functional transactivation assays of these IPF-1 mutant isoforms in a beta-pancreatic tumor cell line transfected with a transcriptional reporter and IPF-1 expression plasmids demonstrate a significant inhibition of basal insulin promoter activity (stronger with the InsCCG243 mutant). We find that the InsCCG243 mutation is linked, in 2 families, to an autosomal dominant-like late-onset form of type 2 diabetes, in which insulin secretion becomes progressively impaired. The lower penetrance D76N and Q59L mutations were more prevalent and were associated with a relative risk of 12.6 for diabetes and with decreased glucose-stimulated insulin-secretion in nondiabetic subjects. We propose that IPF-1 mutations can cause MODY or apparently monogenic late-onset diabetes and that they represent a significant risk factor for type 2 diabetes in humans.
Figures
References
-
- DeFronzo RA. Pathogenesis of type 2 diabetes: metabolic and molecular implications for identifying diabetes genes. Diabet Rev. 1997;5:177–269.
-
- Hanis CL, et al. A genome-wide search for human non–insulin-dependent (type2) diabetes genes reveals a major susceptibility locus on chromosome 2. Nat Genet. 1996;13:161–166. - PubMed
-
- Mahtani MM, et al. Mapping of a gene for type 2 diabetes associated with an insulin secretion defect by a genome scan in Finnish families. Nat Genet. 1996;14:90–94. - PubMed
-
- Hattersley AT. Maturity-onset diabetes of the young: clinical heterogeneity explained by genetic heterogeneity. Diabet Med. 1998;15:15–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
