

The anticonvulsant BW534U87 depresses epileptiform activity in rat hippocampal slices by an adenosine-dependent mechanism and through inhibition of voltage-gated Na+ channels
- PMID: 10556938
- PMCID: PMC1571724
- DOI: 10.1038/sj.bjp.0702881
The anticonvulsant BW534U87 depresses epileptiform activity in rat hippocampal slices by an adenosine-dependent mechanism and through inhibition of voltage-gated Na+ channels
Abstract
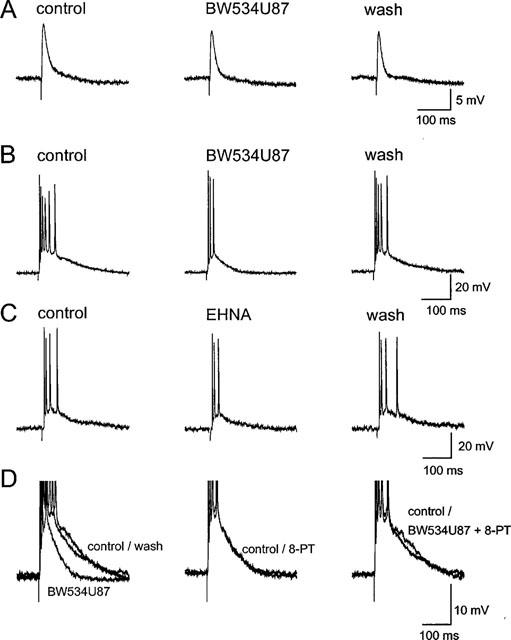
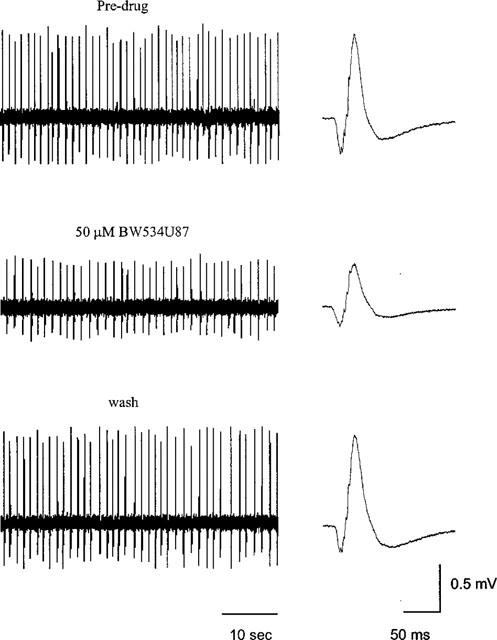
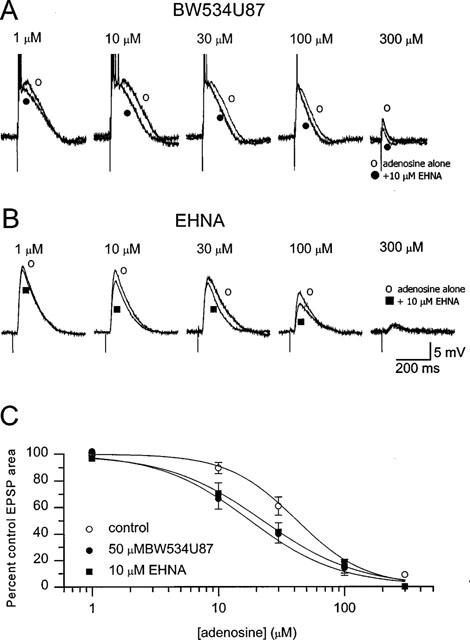
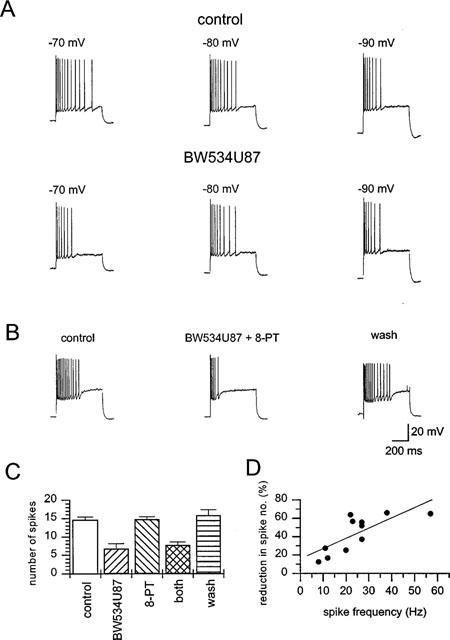
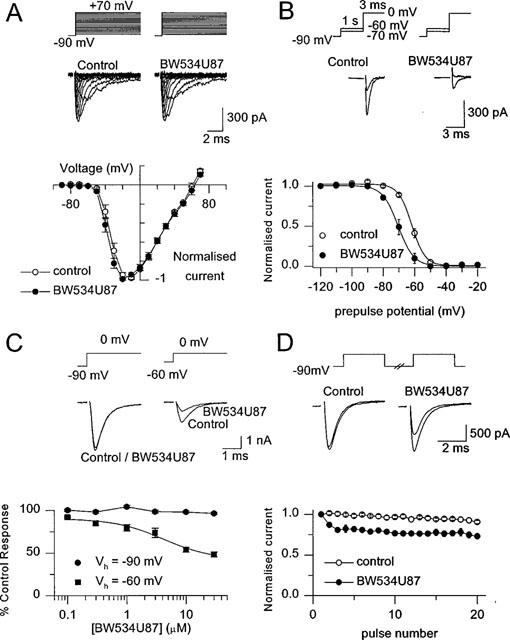
1. The cellular and molecular actions of BW534U87 were studied using intracellular and extracellular recordings from the CA1 region of rat hippocampal slices and whole-cell voltage-clamp recordings of recombinant human brain type IIA Na+ channels expressed in Chinese hamster ovary (CHO) cells. 2. Normal excitatory and inhibitory postsynaptic potentials evoked in hippocampal slices were unaffected by BW534U87 or the adenosine deaminase inhibitor EHNA. However, epileptiform activity was depressed by BW534U87 (50 micronM) and this inhibition was reversed by the adenosine receptor antagonist 8-phenyl theophylline (8-PT, 30 micronM). EHNA (10 micronM) mimicked the effects of BW534U87. Furthermore, BW534U87 enhanced the inhibitory effects of exogenous adenosine on evoked synaptic potentials. BW534U87 (50 micronM) also voltage- and use-dependently inhibited action potentials elicited by current injection, independent of the adenosine system, since it was not affected by 8-PT. 3. In CHO cells expressing the recombinant human brain Na+ channel, BW534U87 produced a concentration- and voltage-dependent inhibition of Na+ currents with a half-maximal inhibitory concentration of 10 micronM at a Vh of -60 mV. Use-dependent inhibition was evident at high-frequencies (20x20 ms pulse train at 10 Hz). 4 In conclusion, BW534U87 blocks hippocampal epileptiform activity by a dual mechanism. The first action is similar to that produced by EHNA and is dependent on endogenous adenosine probably by inhibition of adenosine deaminase. Secondly, BW534U87 directly inhibits voltage-gated Na+ channels in a voltage- and frequency-dependent manner. Both actions of BW534U87 are activity-dependent and may synergistically contribute to its overall anticonvulsant effects in animal models of epilepsy.
Figures
Similar articles
-
Broad spectrum anticonvulsant activity of BW534U87: possible role of an adenosine-dependent mechanism.Pharmacol Biochem Behav. 2002 Dec;74(1):111-8. doi: 10.1016/s0091-3057(02)00956-5. Pharmacol Biochem Behav. 2002. PMID: 12376158
-
Interaction of the antiepileptic drug lamotrigine with recombinant rat brain type IIA Na+ channels and with native Na+ channels in rat hippocampal neurones.Pflugers Arch. 1995 Jul;430(3):437-46. doi: 10.1007/BF00373920. Pflugers Arch. 1995. PMID: 7491269
-
The anticonvulsant actions of sigma receptor ligands in the Mg2+-free model of epileptiform activity in rat hippocampal slices.Br J Pharmacol. 1998 Jul;124(5):917-29. doi: 10.1038/sj.bjp.0701902. Br J Pharmacol. 1998. PMID: 9692777 Free PMC article.
-
The anticonvulsant action of propofol on epileptiform activity in rat hippocampal slices.Anesth Analg. 2004 Oct;99(4):1095-1101. doi: 10.1213/01.ANE.0000130356.22414.2B. Anesth Analg. 2004. PMID: 15385357
-
State-dependent inhibition of Na+ currents by the neuroprotective agent 619C89 in rat hippocampal neurons and in a mammalian cell line expressing rat brain type IIA Na+ channels.Neuroscience. 1996 Aug;73(4):951-62. doi: 10.1016/0306-4522(96)00092-9. Neuroscience. 1996. PMID: 8809814
Cited by
-
Identification of both GABAA receptors and voltage-activated Na(+) channels as molecular targets of anticonvulsant α-asarone.Front Pharmacol. 2014 Mar 11;5:40. doi: 10.3389/fphar.2014.00040. eCollection 2014. Front Pharmacol. 2014. PMID: 24653701 Free PMC article.
-
Endogenous adenosine modulation of 22Na uptake by rat brain synaptosomes.Neurochem Res. 2003 Oct;28(10):1591-5. doi: 10.1023/a:1025638929136. Neurochem Res. 2003. PMID: 14570405
-
Putative Role of Adenosine A1 Receptors in Exogenous Ketone Supplements-Evoked Anti-Epileptic Effect.Int J Mol Sci. 2024 Sep 12;25(18):9869. doi: 10.3390/ijms25189869. Int J Mol Sci. 2024. PMID: 39337356 Free PMC article. Review.
-
Modulators of nucleoside metabolism in the therapy of brain diseases.Curr Top Med Chem. 2011;11(8):1068-86. doi: 10.2174/156802611795347609. Curr Top Med Chem. 2011. PMID: 21401494 Free PMC article. Review.
References
-
- BENDER A.S., WU P.H., PHILLIS J.W. The characterization of [3H] adenosine uptake into rat cerebral cortical synaptosomes. J. Neurochem. 1980;35:629–640. - PubMed
-
- BRUNDEGE J.M., DUNWIDDIE T.V. Role of adenosine as a modulator of synaptic activity in the central nervous system. Adv. Pharmacol. 1997;39:353–391. - PubMed
-
- BURGER R.M., LOWENSTEIN J.M. Preparation and properties of 5′-nucleotidase from smooth muscle of small intestine. J. Biol. Chem. 1970;245:6274–6280. - PubMed
-
- CATTERALL W.A. Cellular and molecular biology of voltage-gated sodium channels. Physiol. Rev. 1992;72:S15–48. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
