

Metabolite Control Overrides Circadian Regulation of Phosphoenolpyruvate Carboxylase Kinase and CO(2) Fixation in Crassulacean Acid Metabolism
- PMID: 10557237
- PMCID: PMC59451
- DOI: 10.1104/pp.121.3.889
Metabolite Control Overrides Circadian Regulation of Phosphoenolpyruvate Carboxylase Kinase and CO(2) Fixation in Crassulacean Acid Metabolism
Abstract
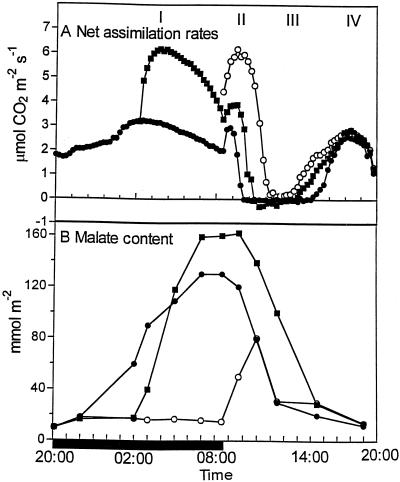
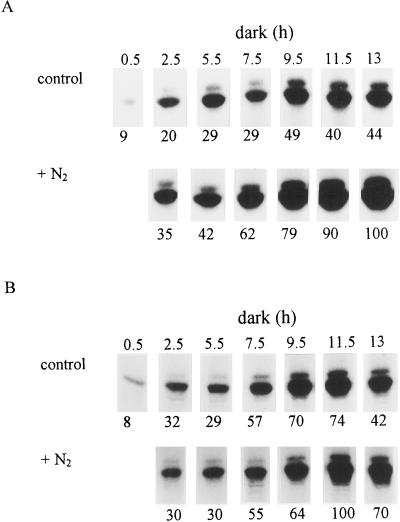
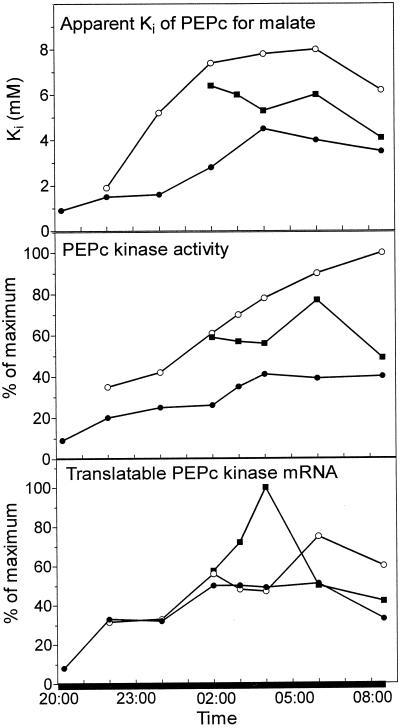
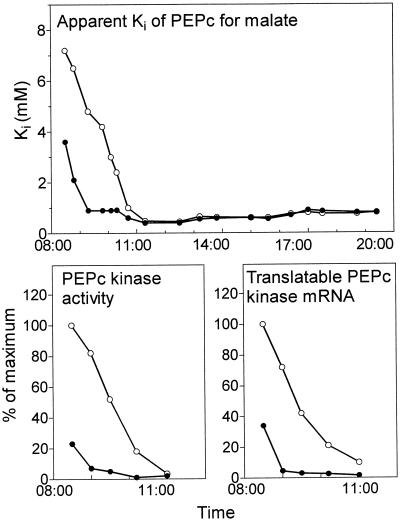
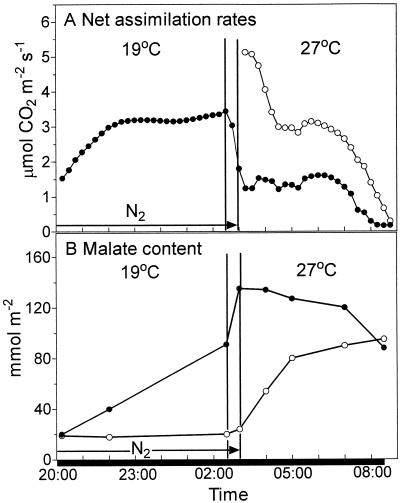
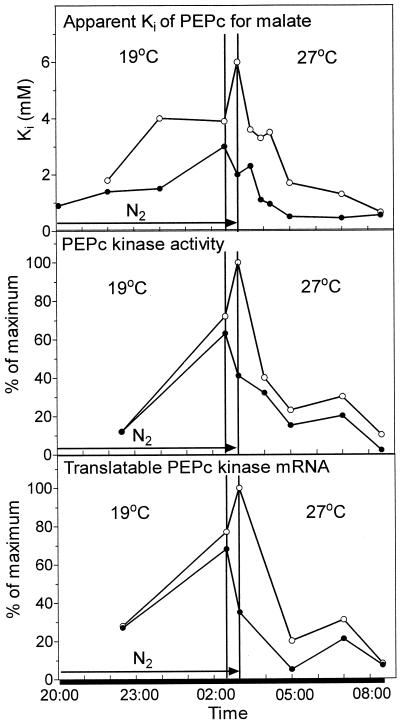
Phosphoenolpyruvate carboxylase (PEPc) catalyzes the primary fixation of CO(2) in Crassulacean acid metabolism plants. Flux through the enzyme is regulated by reversible phosphorylation. PEPc kinase is controlled by changes in the level of its translatable mRNA in response to a circadian rhythm. The physiological significance of changes in the levels of PEPc-kinase-translatable mRNA and the involvement of metabolites in control of the kinase was investigated by subjecting Kalanchoë daigremontiana leaves to anaerobic conditions at night to modulate the magnitude of malate accumulation, or to a rise in temperature at night to increase the efflux of malate from vacuole to cytosol. Changes in CO(2) fixation and PEPc kinase activity reflected those in kinase mRNA. The highest rates of CO(2) fixation and levels of kinase mRNA were observed in leaves subjected to anaerobic treatment for the first half of the night and then transferred to ambient air. In leaves subjected to anaerobic treatment overnight and transferred to ambient air at the start of the day, PEPc-kinase-translatable mRNA and activity, the phosphorylation state of PEPc, and fixation of atmospheric CO(2) were significantly higher than those for control leaves for the first 3 h of the light period. A nighttime temperature increase from 19 degrees C to 27 degrees C led to a rapid reduction in kinase mRNA and activity; however, this was not observed in leaves in which malate accumulation had been prevented by anaerobic treatment. These data are consistent with the hypothesis that a high concentration of malate reduces both kinase mRNA and the accumulation of the kinase itself.
Figures
References
-
- Borland AM, Griffiths H. A comparative study on the regulation of C3 and C4 carboxylation processes in the constitutive Crassulacean acid metabolism (CAM) plant Kalanchoë daigremontiana and the C3-CAM intermediate Clusia minor. Planta. 1997;201:368–378. - PubMed
-
- Fischer A, Kluge M. Studies on carbon flow in Crassulacean acid metabolism during the initial light period. Planta. 1984;160:121–128. - PubMed
-
- Friemert V, Heininger D, Kluge M, Zeigler H. Temperature effects on malic-acid efflux from the vacuoles and on the carboxylation pathways in Crassulacean acid metabolism plants. Planta. 1988;174:453–461. - PubMed
LinkOut - more resources
Full Text Sources
