

Evaluation of functional interaction between K(+) channel alpha- and beta-subunits and putative inactivation gating by Co-expression in Xenopus laevis oocytes
- PMID: 10557249
- PMCID: PMC59464
- DOI: 10.1104/pp.121.3.995
Evaluation of functional interaction between K(+) channel alpha- and beta-subunits and putative inactivation gating by Co-expression in Xenopus laevis oocytes
Abstract
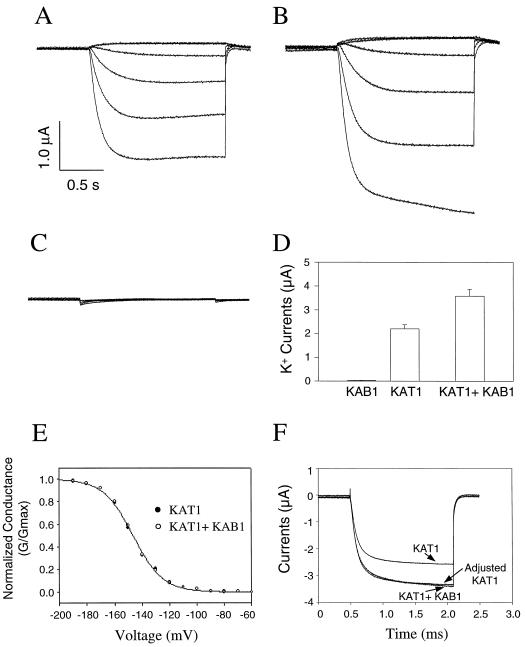
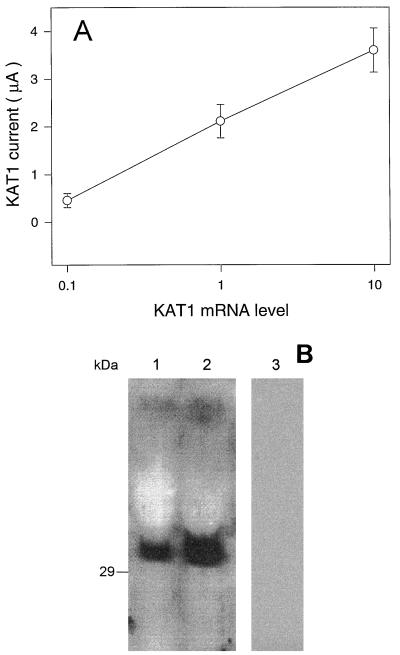
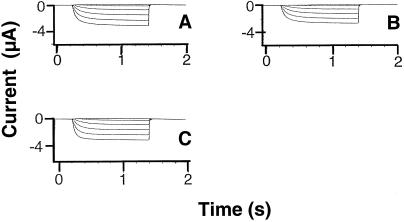
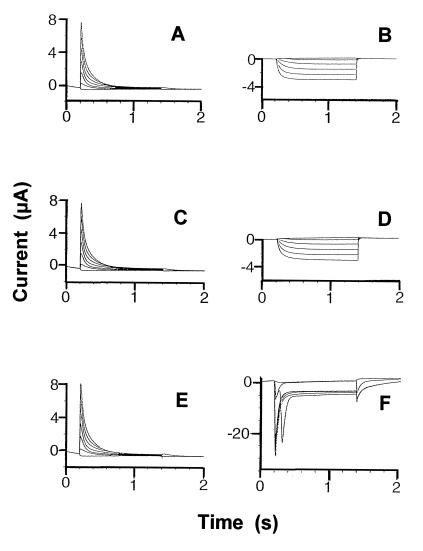
Animal K(+) channel alpha- (pore-forming) subunits form native proteins by association with beta-subunits, which are thought to affect channel function by modifying electrophysiological parameters of currents (often by inducing fast inactivation) or by stabilizing the protein complex. We evaluated the functional association of KAT1, a plant K(+) channel alpha-subunit, and KAB1 (a putative homolog of animal K(+) channel beta-subunits) by co-expression in Xenopus laevis oocytes. Oocytes expressing KAT1 displayed inward-rectifying, non-inactivating K(+) currents that were similar in magnitude to those reported in prior studies. K(+) currents recorded from oocytes expressing both KAT1 and KAB1 had similar gating kinetics. However, co-expression resulted in greater total current, consistent with the possibility that KAB1 is a beta-subunit that stabilizes and therefore enhances surface expression of K(+) channel protein complexes formed by alpha-subunits such as KAT1. K(+) channel protein complexes formed by alpha-subunits such as KAT1 that undergo (voltage-dependent) inactivation do so by means of a "ball and chain" mechanism; the ball portion of the protein complex (which can be formed by the N terminus of either an alpha- or beta-subunit) occludes the channel pore. KAT1 was co-expressed in oocytes with an animal K(+) channel alpha-subunit (hKv1.4) known to contain the N-terminal ball and chain. Inward currents through heteromeric hKv1. 4:KAT1 channels did undergo typical voltage-dependent inactivation. These results suggest that inward currents through K(+) channel proteins formed at least in part by KAT1 polypeptides are capable of inactivation, but the structural component facilitating inactivation is not present when channel complexes are formed by either KAT1 or KAB1 in the absence of additional subunits.
Figures
Similar articles
-
Co-expression of calcium-dependent protein kinase with the inward rectified guard cell K+ channel KAT1 alters current parameters in Xenopus laevis oocytes.Plant Cell Physiol. 2000 Jun;41(6):785-90. doi: 10.1093/pcp/41.6.785. Plant Cell Physiol. 2000. PMID: 10945349
-
Physical association of KAB1 with plant K+ channel alpha subunits.Plant Cell. 1996 Sep;8(9):1545-53. doi: 10.1105/tpc.8.9.1545. Plant Cell. 1996. PMID: 8837508 Free PMC article.
-
The N-terminus of the K channel KAT1 controls its voltage-dependent gating by altering the membrane electric field.Biophys J. 1998 Jun;74(6):2953-62. doi: 10.1016/S0006-3495(98)78002-6. Biophys J. 1998. PMID: 9635749 Free PMC article.
-
Inward rectifier potassium channels. Cloning, expression and structure-function studies.Jpn Heart J. 1996 Sep;37(5):651-60. doi: 10.1536/ihj.37.651. Jpn Heart J. 1996. PMID: 8973378 Review.
-
Functional and molecular aspects of voltage-gated K+ channel beta subunits.Ann N Y Acad Sci. 1999 Apr 30;868:344-55. doi: 10.1111/j.1749-6632.1999.tb11296.x. Ann N Y Acad Sci. 1999. PMID: 10414304 Review.
Cited by
-
Interactions between plasma membrane aquaporins modulate their water channel activity.Plant Cell. 2004 Jan;16(1):215-28. doi: 10.1105/tpc.017194. Epub 2003 Dec 11. Plant Cell. 2004. PMID: 14671024 Free PMC article.
-
Physical and functional interaction of the Arabidopsis K(+) channel AKT2 and phosphatase AtPP2CA.Plant Cell. 2002 May;14(5):1133-46. doi: 10.1105/tpc.000943. Plant Cell. 2002. PMID: 12034902 Free PMC article.
-
AtKC1, a silent Arabidopsis potassium channel alpha -subunit modulates root hair K+ influx.Proc Natl Acad Sci U S A. 2002 Mar 19;99(6):4079-84. doi: 10.1073/pnas.052677799. Proc Natl Acad Sci U S A. 2002. PMID: 11904452 Free PMC article.
-
Genome-wide identification of kiwifruit K+ channel Shaker family members and their response to low-K+ stress.BMC Plant Biol. 2024 Sep 6;24(1):833. doi: 10.1186/s12870-024-05555-x. BMC Plant Biol. 2024. PMID: 39243055 Free PMC article.
-
Ectopic expression of the K+ channel β subunits from Puccinellia tenuiflora (KPutB1) and rice (KOB1) alters K+ homeostasis of yeast and Arabidopsis.Mol Biotechnol. 2011 May;48(1):76-86. doi: 10.1007/s12033-010-9349-3. Mol Biotechnol. 2011. PMID: 21108023
References
-
- Cao Y, Crawford NM, Schroeder JI. Amino terminus and the first four membrane-spanning segments of the Arabidopsis K+ channel KAT1 confer inward-rectification property of plant-animal chimeric channels. J Biol Chem. 1995;270:17697–17701. - PubMed
-
- Dolly JO, Rettig J, Scott VES, Parcej DN, Wittka R, Sewing S, Pongs O. Oligomeric and subunit structures of neuronal voltage-sensitive K+ channels. Biochem Soc Trans. 1994;22:473–478. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
