

c-Myc enhances protein synthesis and cell size during B lymphocyte development
- PMID: 10557294
- PMCID: PMC23921
- DOI: 10.1073/pnas.96.23.13180
c-Myc enhances protein synthesis and cell size during B lymphocyte development
Abstract
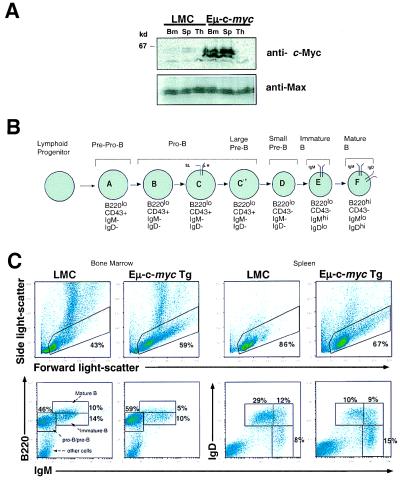
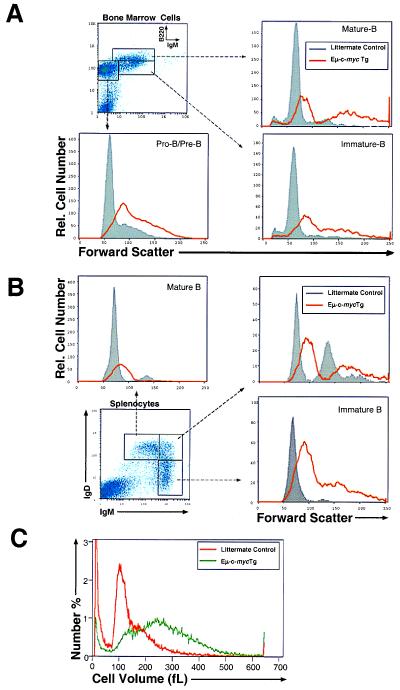
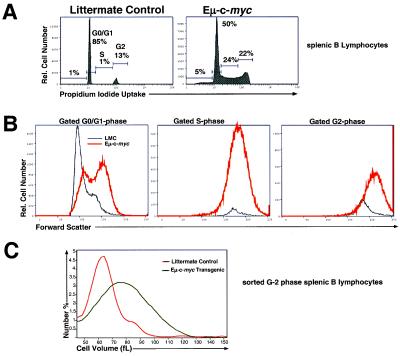
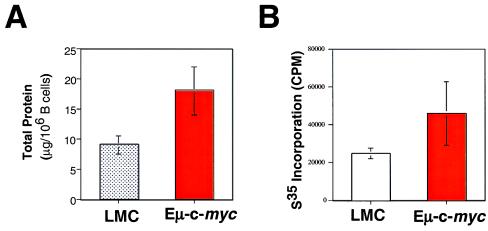
Members of the myc family of nuclear protooncogenes play roles in cell proliferation, differentiation, and apoptosis. Moreover, inappropriate expression of c-myc genes contributes to the development of many types of cancers, including B cell lymphomas in humans. Although Myc proteins have been shown to function as transcription factors, their immediate effects on the cell have not been well defined. Here we have utilized a murine model of lymphomagenesis (Emu-myc mice) to show that constitutive expression of a c-myc transgene under control of the Ig heavy-chain enhancer (Emu) results in an increase in cell size of normal pretransformed B lymphocytes at all stages of B cell development. Furthermore, we show that c-Myc-induced growth occurs independently of cell cycle phase and correlates with an increase in protein synthesis. These results suggest that Myc may normally function by coordinating expression of growth-related genes in response to mitogenic signals. Deregulated c-myc expression may predispose to cancer by enhancing cell growth to levels required for unrestrained cell division.
Figures
References
-
- Killander D, Zetterberg A. Exp Cell Res. 1965;38:272–284. - PubMed
-
- Neufeld T P, Edgar B A. Curr Opin Cell Biol. 1998;10:784–790. - PubMed
-
- Blackwood E M, Eisenman R N. Science. 1991;251:1211–1217. - PubMed
-
- Prendergast G C, Lawe D, Ziff E B. Cell. 1991;65:395–407. - PubMed
-
- Kretzner L, Blackwood E M, Eisenman R N. Nature (London) 1992;359:426–429. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
