

Morphological analysis of the mammalian postcranium: a developmental perspective
- PMID: 10557306
- PMCID: PMC23933
- DOI: 10.1073/pnas.96.23.13247
Morphological analysis of the mammalian postcranium: a developmental perspective
Abstract
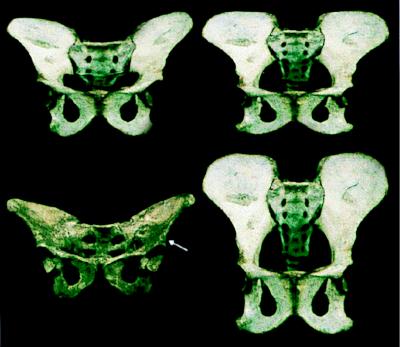
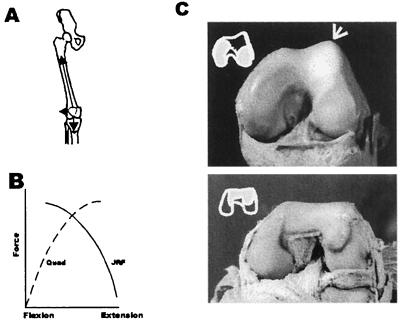
The past two decades have greatly improved our knowledge of vertebrate skeletal morphogenesis. It is now clear that bony morphology lacks individual descriptive specification and instead results from an interplay between positional information assigned during early limb bud deployment and its "execution" by highly conserved cellular response programs of derived connective tissue cells (e.g., chondroblasts and osteoblasts). Selection must therefore act on positional information and its apportionment, rather than on more individuated aspects of presumptive adult morphology. We suggest a trait classification system that can help integrate these findings in both functional and phylogenetic examinations of fossil mammals and provide examples from the human fossil record.
Figures
Similar articles
-
Developmental mechanisms of vertebrate limb evolution.Novartis Found Symp. 2001;232:47-57; discussion 57-62. doi: 10.1002/0470846658.ch5. Novartis Found Symp. 2001. PMID: 11277086 Review.
-
Development and evolution of the muscles of the pelvic fin.PLoS Biol. 2011 Oct;9(10):e1001168. doi: 10.1371/journal.pbio.1001168. Epub 2011 Oct 4. PLoS Biol. 2011. PMID: 21990962 Free PMC article.
-
Variation in mouse pelvic morphology maps to locations enriched in Sox9 Class II and Pitx1 regulatory features.J Exp Zool B Mol Dev Evol. 2020 Mar;334(2):100-112. doi: 10.1002/jez.b.22926. Epub 2020 Feb 3. J Exp Zool B Mol Dev Evol. 2020. PMID: 32017444
-
Locomotion in some small to medium-sized mammals: a geometric morphometric analysis of the penultimate lumbar vertebra, pelvis and hindlimbs.Zoology (Jena). 2013 Dec;116(6):356-71. doi: 10.1016/j.zool.2013.08.007. Epub 2013 Oct 5. Zoology (Jena). 2013. PMID: 24182890
-
Endochondral ossification and the evolution of limb proportions.Wiley Interdiscip Rev Dev Biol. 2020 Jul;9(4):e373. doi: 10.1002/wdev.373. Epub 2020 Jan 29. Wiley Interdiscip Rev Dev Biol. 2020. PMID: 31997553 Review.
Cited by
-
Evolution of the female pelvis and relationships to pelvic organ prolapse.Int Urogynecol J Pelvic Floor Dysfunct. 2005 Jul-Aug;16(4):315-20. doi: 10.1007/s00192-004-1258-1. Epub 2005 Jan 15. Int Urogynecol J Pelvic Floor Dysfunct. 2005. PMID: 15654501 Review.
-
Ontogenetic changes in the internal and external morphology of the ilium in modern humans.J Anat. 2011 Mar;218(3):324-35. doi: 10.1111/j.1469-7580.2011.01342.x. J Anat. 2011. PMID: 21323915 Free PMC article.
-
The shape of the Neandertal femur is primarily the consequence of a hyperpolar body form.Proc Natl Acad Sci U S A. 2003 Jun 10;100(12):6926-9. doi: 10.1073/pnas.1232340100. Epub 2003 May 21. Proc Natl Acad Sci U S A. 2003. PMID: 12761384 Free PMC article.
-
How reliable are human phylogenetic hypotheses?Proc Natl Acad Sci U S A. 2000 Apr 25;97(9):5003-6. doi: 10.1073/pnas.97.9.5003. Proc Natl Acad Sci U S A. 2000. PMID: 10781112 Free PMC article.
-
Metacarpal torsion in apes, humans, and early Australopithecus: implications for manipulatory abilities.PeerJ. 2015 Oct 6;3:e1311. doi: 10.7717/peerj.1311. eCollection 2015. PeerJ. 2015. PMID: 26500820 Free PMC article.
References
-
- Hillis D M, Huselsenbeck J P, Cunningham C W. Science. 1994;264:671–677. - PubMed
-
- Douzery E, Catzeflis F M. J Mol Evol. 1995;41:622–636. - PubMed
-
- Krings M, Stake A, Schmitz R W, Krainitzki H, Stoneking M, Paabo S. Cell. 1997;90:19–30. - PubMed
-
- Dobzhansky T. Am Nat. 1956;40:337–347.
-
- Gould S J, Lewontin R C. Proc R Soc London Ser B. 1979;205:147–164. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
