

Thickness and elasticity of gram-negative murein sacculi measured by atomic force microscopy
- PMID: 10559150
- PMCID: PMC94159
- DOI: 10.1128/JB.181.22.6865-6875.1999
Thickness and elasticity of gram-negative murein sacculi measured by atomic force microscopy
Abstract
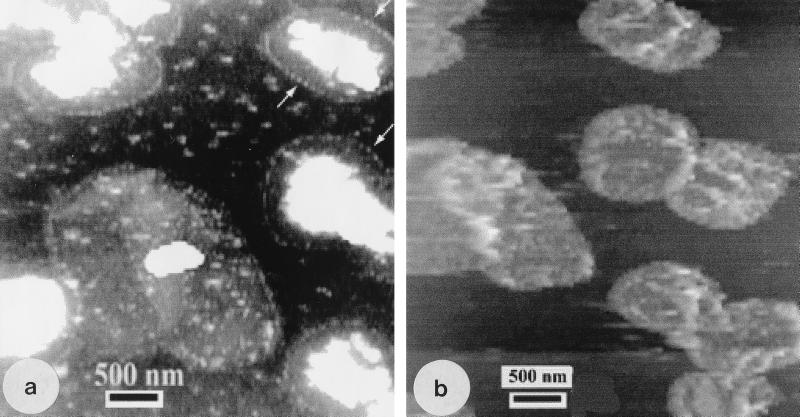
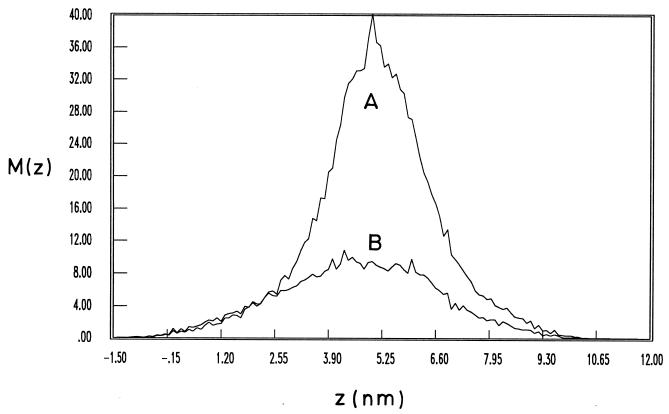
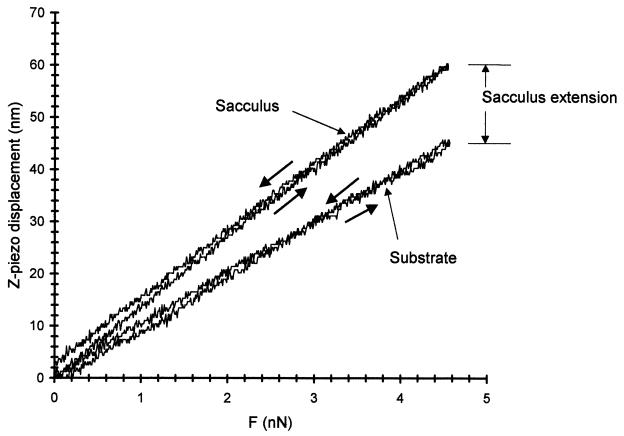
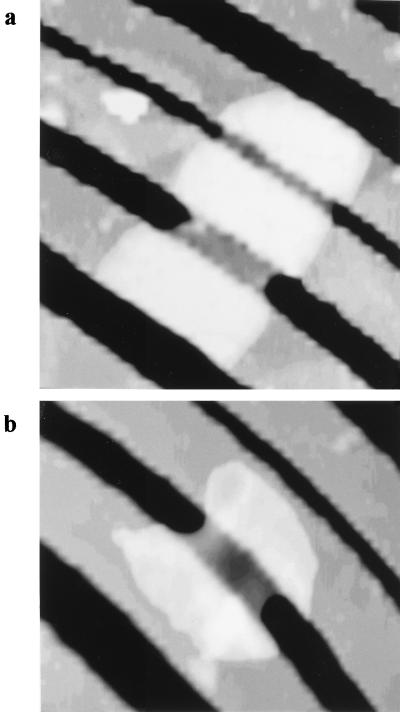
Atomic force microscopy was used to measure the thickness of air-dried, collapsed murein sacculi from Escherichia coli K-12 and Pseudomonas aeruginosa PAO1. Air-dried sacculi from E. coli had a thickness of 3.0 nm, whereas those from P. aeruginosa were 1.5 nm thick. When rehydrated, the sacculi of both bacteria swelled to double their anhydrous thickness. Computer simulation of a section of a model single-layer peptidoglycan network in an aqueous solution with a Debye shielding length of 0.3 nm gave a mass distribution full width at half height of 2.4 nm, in essential agreement with these results. When E. coli sacculi were suspended over a narrow groove that had been etched into a silicon surface and the tip of the atomic force microscope used to depress and stretch the peptidoglycan, an elastic modulus of 2.5 x 10(7) N/m(2) was determined for hydrated sacculi; they were perfectly elastic, springing back to their original position when the tip was removed. Dried sacculi were more rigid with a modulus of 3 x 10(8) to 4 x 10(8) N/m(2) and at times could be broken by the atomic force microscope tip. Sacculi aligned over the groove with their long axis at right angles to the channel axis were more deformable than those with their long axis parallel to the groove axis, as would be expected if the peptidoglycan strands in the sacculus were oriented at right angles to the long cell axis of this gram-negative rod. Polar caps were not found to be more rigid structures but collapsed to the same thickness as the cylindrical portions of the sacculi. The elasticity of intact E. coli sacculi is such that, if the peptidoglycan strands are aligned in unison, the interstrand spacing should increase by 12% with every 1 atm increase in (turgor) pressure. Assuming an unstressed hydrated interstrand spacing of 1.3 nm (R. E. Burge, A. G. Fowler, and D. A. Reaveley, J. Mol. Biol. 117:927-953, 1977) and an internal turgor pressure of 3 to 5 atm (or 304 to 507 kPa) (A. L. Koch, Adv. Microbial Physiol. 24:301-366, 1983), the natural interstrand spacing in cells would be 1.6 to 2.0 nm. Clearly, if large macromolecules of a diameter greater than these spacings are secreted through this layer, the local ordering of the peptidoglycan must somehow be disrupted.
Figures
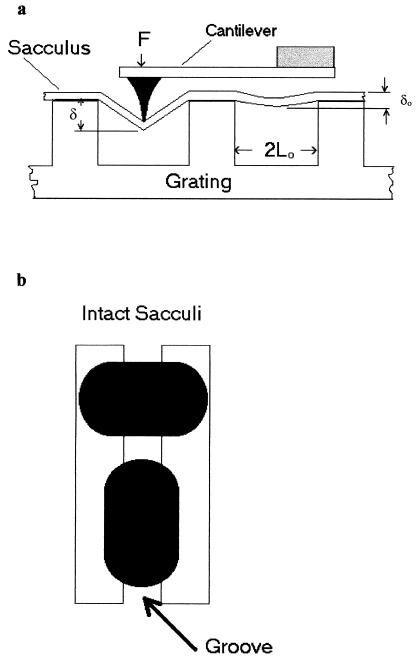
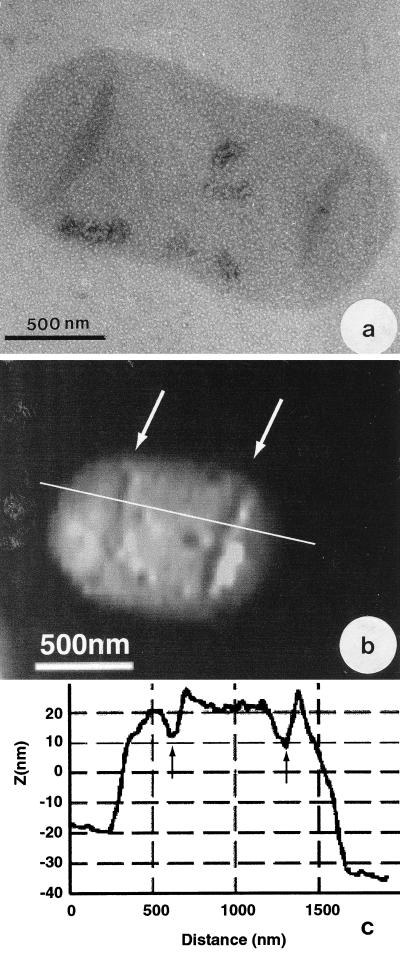

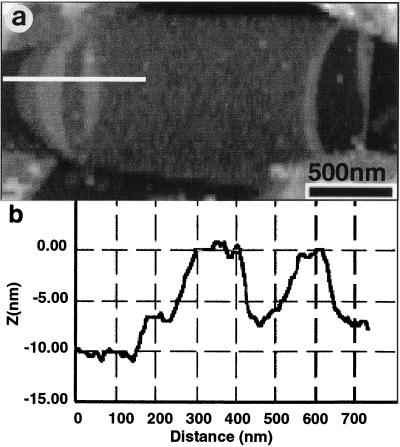
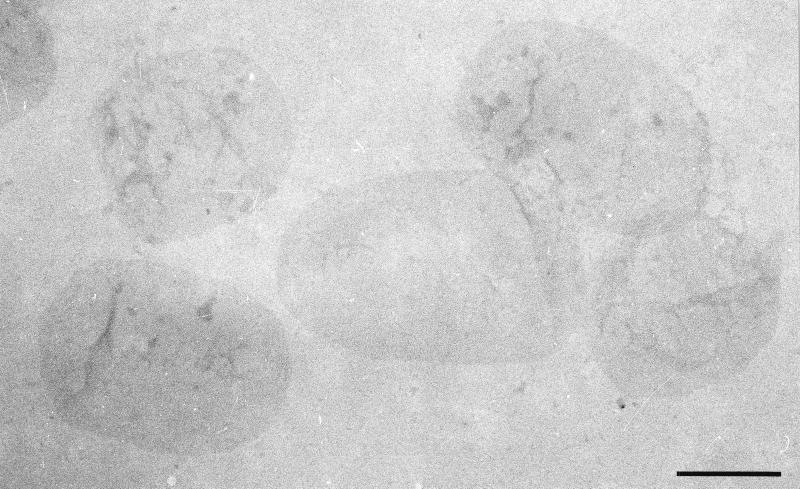
References
-
- Barnickel G, Naumann D, Bradczek H, Labischinski H, Giesbrecht P. Computer aided molecular modeling of the three-dimensional structure of bacterial peptidoglycan. In: Hakenbeck R, Höltje J-V, Labischinski H, editors. The target of penicillin. Berlin, Germany: Walter de Gruyter & Co.; 1983. pp. 61–66.
-
- Bayer M E. Zones of membrane adhesion in the cryofixed envelope of Escherichia coli. J Struct Biol. 1991;197:268–280. - PubMed
-
- Beveridge T J. The periplasmic space and the concept of the periplasm in gram-positive and gram-negative bacteria. ASM News. 1995;61:125–130.
-
- Beveridge T J. Ultrastructure, chemistry, and function of the bacterial wall. Int Rev Cytol. 1981;72:229–317. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
