

Partition of the linear plasmid N15: interactions of N15 partition functions with the sop locus of the F plasmid
- PMID: 10559154
- PMCID: PMC94163
- DOI: 10.1128/JB.181.22.6898-6906.1999
Partition of the linear plasmid N15: interactions of N15 partition functions with the sop locus of the F plasmid
Abstract
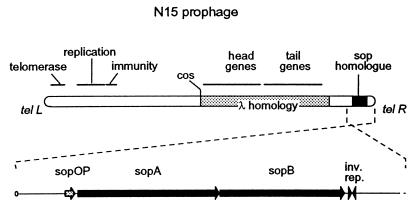
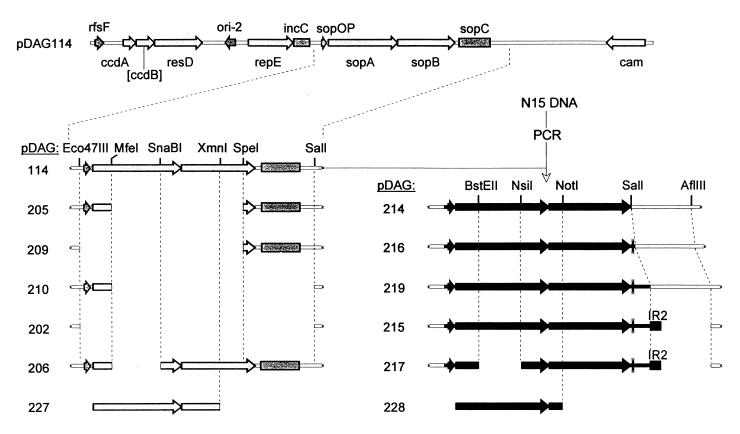
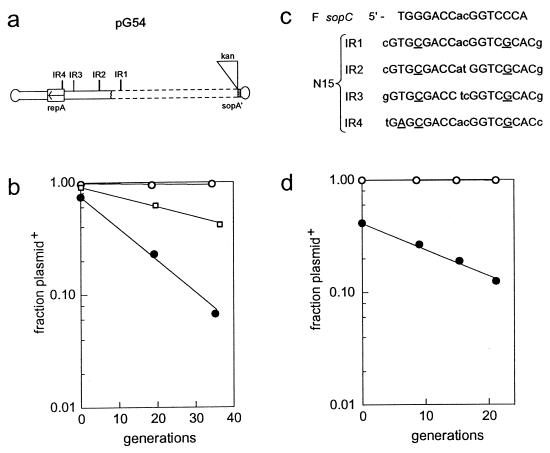
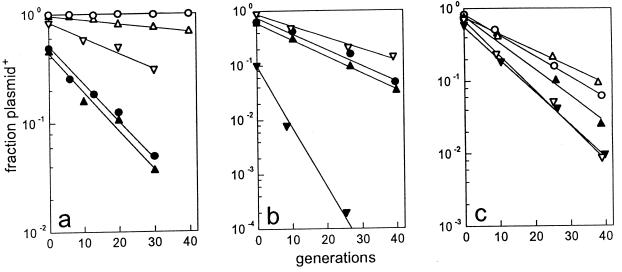
A locus close to one end of the linear N15 prophage closely resembles the sop operon which governs partition of the F plasmid; the promoter region contains similar operator sites, and the two putative gene products have extensive amino acid identity with the SopA and -B proteins of F. Our aim was to ascertain whether the N15 sop homologue functions in partition, to identify the centromere site, and to examine possible interchangeability of function with the F Sop system. When expressed at a moderate level, N15 SopA and -B proteins partly stabilize mini-F which lacks its own sop operon but retains the sopC centromere. The stabilization does not depend on increased copy number. Likewise, an N15 mutant with most of its sop operon deleted is partly stabilized by F Sop proteins and fully stabilized by its own. Four inverted repeat sequences similar to those of sopC were located in N15. They are distant from the sop operon and from each other. Two of these were shown to stabilize a mini-F sop deletion mutant when N15 Sop proteins were provided. Provision of the SopA homologue to plasmids with a sopA deletion resulted in further destabilization of the plasmid. The N15 Sop proteins exert effective, but incomplete, repression at the F sop promoter. We conclude that the N15 sop locus determines stable inheritance of the prophage by using dispersed centromere sites. The SopB-centromere and SopA-operator interactions show partial functional overlap between N15 and F. SopA of each plasmid appears to interact with SopB of the other, but in a way that is detrimental to plasmid maintenance.
Figures



Similar articles
-
Mapping of functional domains in F plasmid partition proteins reveals a bipartite SopB-recognition domain in SopA.J Mol Biol. 2003 Jun 20;329(5):875-89. doi: 10.1016/s0022-2836(03)00525-4. J Mol Biol. 2003. PMID: 12798679
-
[Sop proteins can cause transcriptional silencing of genes located close to the centromere sites of linear plasmid N15].Mol Biol (Mosk). 2010 Mar-Apr;44(2):294-300. Mol Biol (Mosk). 2010. PMID: 20586190 Russian.
-
Extended function of plasmid partition genes: the Sop system of linear phage-plasmid N15 facilitates late gene expression.J Bacteriol. 2008 May;190(10):3538-45. doi: 10.1128/JB.01993-07. Epub 2008 Mar 21. J Bacteriol. 2008. PMID: 18359814 Free PMC article.
-
Replication and Maintenance of Linear Phage-Plasmid N15.Microbiol Spectr. 2015 Feb;3(1):PLAS-0032-2014. doi: 10.1128/microbiolspec.PLAS-0032-2014. Microbiol Spectr. 2015. PMID: 26104561 Review.
-
N15: the linear phage-plasmid.Plasmid. 2011 Mar;65(2):102-9. doi: 10.1016/j.plasmid.2010.12.004. Epub 2010 Dec 23. Plasmid. 2011. PMID: 21185326 Review.
Cited by
-
Segrosome Complex Formation during DNA Trafficking in Bacterial Cell Division.Front Mol Biosci. 2016 Sep 9;3:51. doi: 10.3389/fmolb.2016.00051. eCollection 2016. Front Mol Biosci. 2016. PMID: 27668216 Free PMC article. Review.
-
Addressing the role of centromere sites in activation of ParB proteins for partition complex assembly.PLoS One. 2020 May 7;15(5):e0226472. doi: 10.1371/journal.pone.0226472. eCollection 2020. PLoS One. 2020. PMID: 32379828 Free PMC article.
-
Characterization of the partitioning system of Myxococcus plasmid pMF1.PLoS One. 2011;6(12):e28122. doi: 10.1371/journal.pone.0028122. Epub 2011 Dec 9. PLoS One. 2011. PMID: 22174771 Free PMC article.
-
Identification of a Potential Membrane-Targeting Sequence in the C-Terminus of the F Plasmid Segregation Protein SopA.J Membr Biol. 2021 Jun;254(3):243-257. doi: 10.1007/s00232-020-00157-8. Epub 2021 Jan 11. J Membr Biol. 2021. PMID: 33427942
-
Comprehensive Genomic Analysis of Klebsiella pneumoniae and Its Temperate N-15-like Phage: From Isolation to Functional Annotation.Microorganisms. 2025 Apr 15;13(4):908. doi: 10.3390/microorganisms13040908. Microorganisms. 2025. PMID: 40284744 Free PMC article.
References
-
- Austin S, Wierzbecki A. Two mini-F encoded proteins are essential for equipartition. Plasmid. 1983;10:73–81. - PubMed
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Current protocols in molecular biology. New York, N.Y: Wiley-Interscience; 1989.
-
- Casadaban M J, Cohen S N. Analysis of gene control signals by DNA fusion and cloning in Escherichia coli. J Mol Biol. 1980;138:179–207. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
