

Differential methylation of Epstein-Barr virus latency promoters facilitates viral persistence in healthy seropositive individuals
- PMID: 10559309
- PMCID: PMC113046
- DOI: 10.1128/JVI.73.12.9959-9968.1999
Differential methylation of Epstein-Barr virus latency promoters facilitates viral persistence in healthy seropositive individuals
Abstract
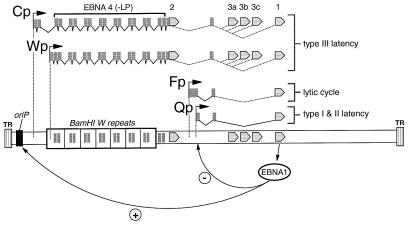
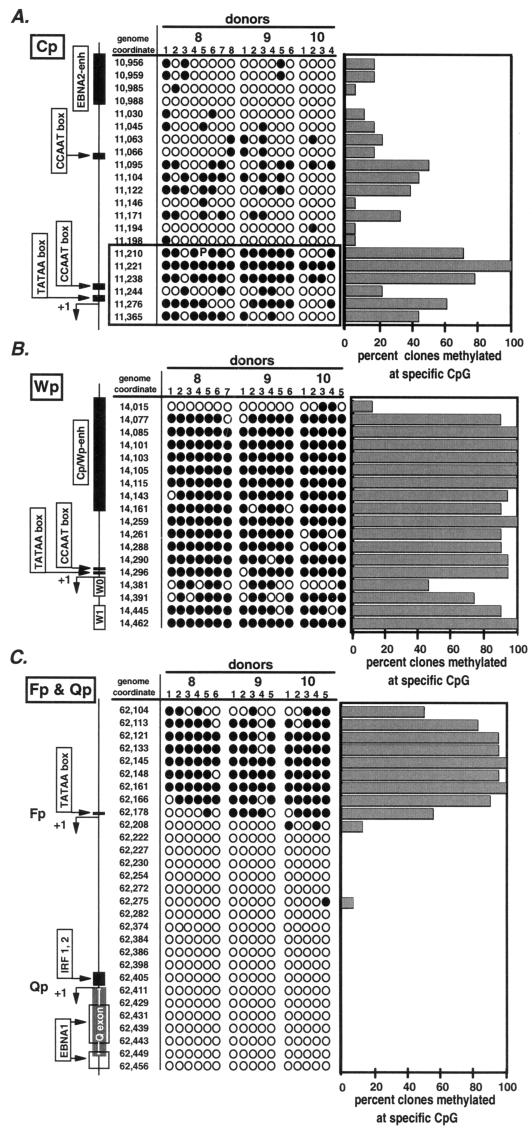
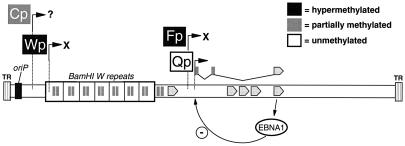
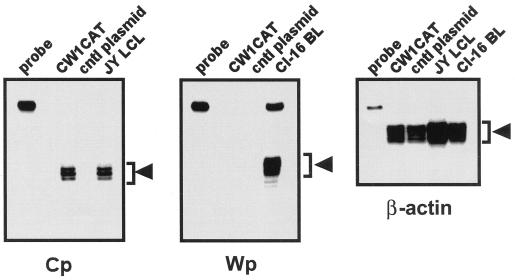
Epstein-Barr virus (EBV) establishes a life-long infection in humans, with distinct viral latency programs predominating during acute and chronic phases of infection. Only a subset of the EBV latency-associated antigens present during the acute phase of EBV infection are expressed in the latently infected memory B cells that serve as the long-term EBV reservoir. Since the EBV immortalization program elicits a potent cellular immune response, downregulation of viral gene expression in the long-term latency reservoir is likely to facilitate evasion of the immune response and persistence of EBV in the immunocompetent host. Tissue culture and tumor models of restricted EBV latency have consistently demonstrated a critical role for methylation of the viral genome in maintaining the restricted pattern of latency-associated gene expression. Here we extend these observations to demonstrate that the EBV genomes in the memory B-cell reservoir are also heavily and discretely methylated. This analysis reveals that methylation of the viral genome is a normal aspect of EBV infection in healthy immunocompetent individuals and is not restricted to the development of EBV-associated tumors. In addition, the pattern of methylation very likely accounts for the observed inhibition of the EBV immortalization program and the establishment and maintenance of a restricted latency program. Thus, EBV appears to be the first example of a parasite that usurps the host cell-directed methylation system to regulate pathogen gene expression and thereby establish a chronic infection.
Figures
Similar articles
-
CpG methylation of the major Epstein-Barr virus latency promoter in Burkitt's lymphoma and Hodgkin's disease.Blood. 1996 Oct 15;88(8):3129-36. Blood. 1996. PMID: 8874213
-
Host-cell-determined methylation of specific Epstein-Barr virus promoters regulates the choice between distinct viral latency programs.Mol Cell Biol. 1997 Jan;17(1):364-77. doi: 10.1128/MCB.17.1.364. Mol Cell Biol. 1997. PMID: 8972217 Free PMC article.
-
Methylation of the EBV genome and establishment of restricted latency in low-passage EBV-infected 293 epithelial cells.Virology. 2002 Jul 20;299(1):109-21. doi: 10.1006/viro.2002.1457. Virology. 2002. PMID: 12167346
-
Epstein-Barr virus immortalization and latency.Front Biosci. 1999 Mar 15;4:D346-71. doi: 10.2741/rowe. Front Biosci. 1999. PMID: 10077545 Review.
-
Epigenotypes of latent herpesvirus genomes.Curr Top Microbiol Immunol. 2006;310:61-80. doi: 10.1007/3-540-31181-5_5. Curr Top Microbiol Immunol. 2006. PMID: 16909907 Review.
Cited by
-
Mining for viral fragments in methylation enriched sequencing data.Front Genet. 2015 Feb 4;6:16. doi: 10.3389/fgene.2015.00016. eCollection 2015. Front Genet. 2015. PMID: 25699076 Free PMC article.
-
Reactivation of the human cytomegalovirus major immediate-early regulatory region and viral replication in embryonal NTera2 cells: role of trichostatin A, retinoic acid, and deletion of the 21-base-pair repeats and modulator.J Virol. 2001 Feb;75(4):1581-93. doi: 10.1128/JVI.75.4.1581-1593.2001. J Virol. 2001. PMID: 11160656 Free PMC article.
-
Protein-DNA binding and CpG methylation at nucleotide resolution of latency-associated promoters Qp, Cp, and LMP1p of Epstein-Barr virus.J Virol. 2001 Mar;75(6):2584-96. doi: 10.1128/JVI.75.6.2584-2596.2001. J Virol. 2001. PMID: 11222681 Free PMC article.
-
EBV latency types adopt alternative chromatin conformations.PLoS Pathog. 2011 Jul;7(7):e1002180. doi: 10.1371/journal.ppat.1002180. Epub 2011 Jul 28. PLoS Pathog. 2011. PMID: 21829357 Free PMC article.
-
The Role of Gammaherpesviruses in Cancer Pathogenesis.Pathogens. 2016 Feb 6;5(1):18. doi: 10.3390/pathogens5010018. Pathogens. 2016. PMID: 26861404 Free PMC article. Review.
References
-
- Babcock G J, Decker L L, Volk M, Thorley-Lawson D A. EBV persistence in memory B cells in vivo. Immunity. 1998;9:395–404. - PubMed
-
- Barlow D P. Methylation and imprinting: from host defense to gene regulation? Science. 1993;260:309–310. - PubMed
-
- Bender J. Cytosine methylation of repeated sequences in eukaryotes: the role of DNA pairing. Trends Biochem Sci. 1998;23:252–256. - PubMed
-
- Bestor T H. Gene silencing. Methylation meets acetylation. Nature. 1998;393:311–312. . (News; comment.) - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
