

Mutational analysis of bovine viral diarrhea virus RNA-dependent RNA polymerase
- PMID: 10559328
- PMCID: PMC113065
- DOI: 10.1128/JVI.73.12.10129-10136.1999
Mutational analysis of bovine viral diarrhea virus RNA-dependent RNA polymerase
Abstract
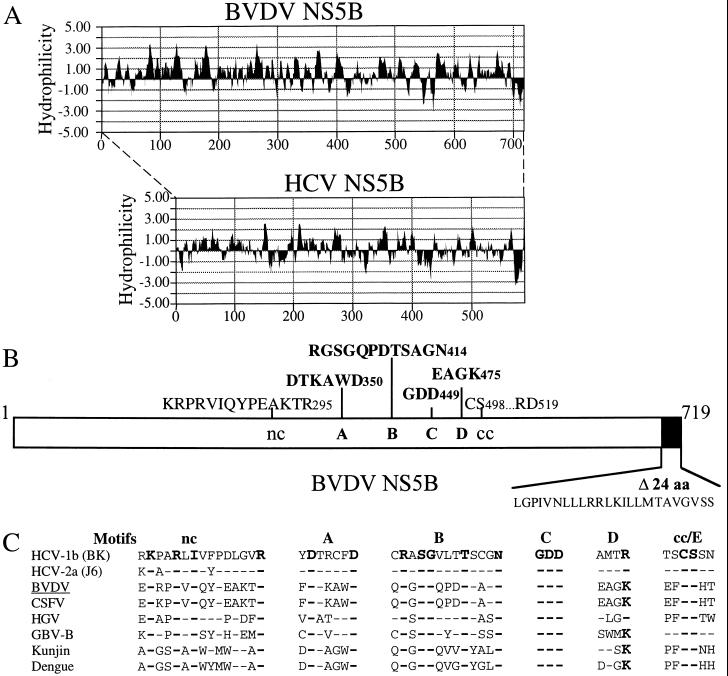
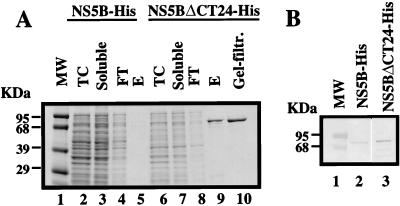
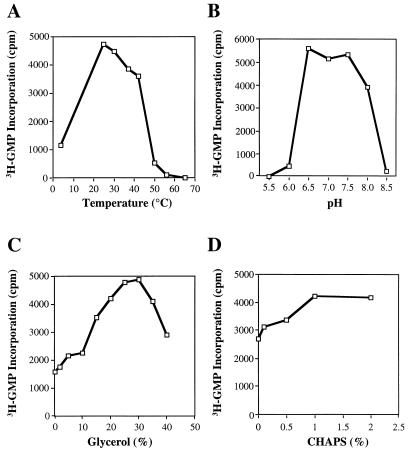
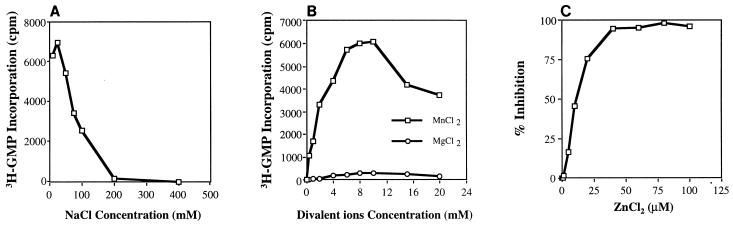
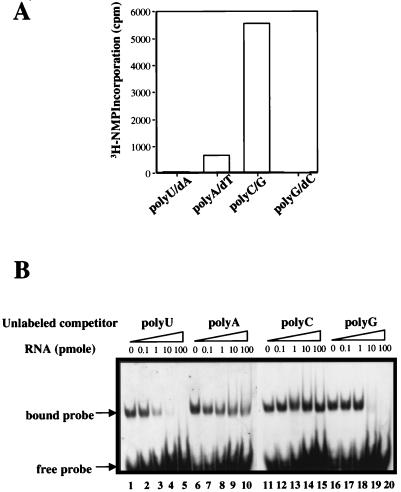
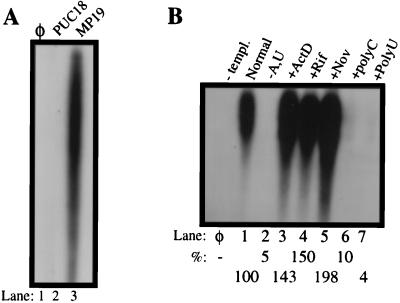
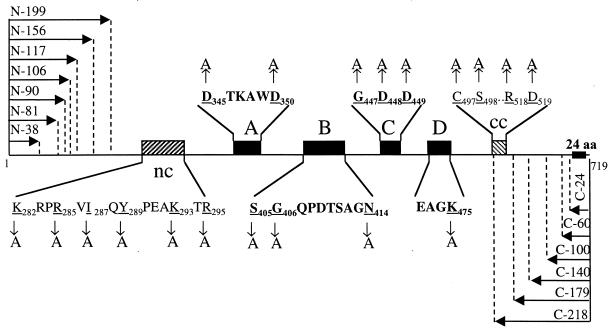
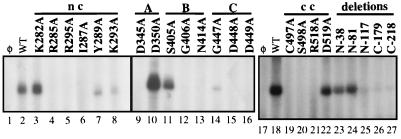
Recombinant bovine viral diarrhea virus (BVDV) nonstructural protein 5B (NS5B) produced in insect cells has been shown to possess an RNA-dependent RNA polymerase (RdRp) activity. Our initial attempt to produce the full-length BVDV NS5B with a C-terminal hexahistidine tag in Escherichia coli failed due to the expression of insoluble products. Prompted by a recent report that removal of the C-terminal hydrophobic domain significantly improved the solubility of hepatitis C virus (HCV) NS5B, we constructed a similar deletion of 24 amino acids at the C terminus of BVDV NS5B. The resulting fusion protein, NS5BDeltaCT24-His, was purified to homogeneity and demonstrated to direct RNA replication via both primer-dependent (elongative) and primer-independent (de novo) mechanisms. Furthermore, BVDV RdRp was found to utilize a circular single-stranded DNA as a template for RNA synthesis, suggesting that synthesis does not require ends in the template. In addition to the previously described polymerase motifs A, B, C, and D, alignments with other flavivirus sequences revealed two additional motifs, one N-terminal to motif A and one C-terminal to motif D. Extensive alanine substitutions showed that while most mutations had similar effects on both elongative and de novo RNA syntheses, some had selective effects. Finally, deletions of up to 90 amino acids from the N terminus did not significantly affect RdRp activities, whereas deletions of more than 24 amino acids at the C terminus resulted in either insoluble products or soluble proteins (DeltaCT179 and DeltaCT218) that lacked RdRp activities.
Figures
References
-
- Adkins S, Quadt R, Choi T J, Ahlquist P, German T. An RNA-dependent RNA polymerase activity associated with virions of tomato spotted wilt virus, a plant- and insect-infecting bunyavirus. Virology. 1995;207:308–311. - PubMed
-
- Chon S K, Perez D R, Donis R O. Genetic analysis of the internal ribosome entry segment of bovine viral diarrhea virus. Virology. 1998;251:370–381. - PubMed
-
- Collett M S, Larson R, Gold C, Strick D, Anderson D K, Purchio A F. Molecular cloning and nucleotide sequence of the pestivirus bovine viral diarrhea virus. Virology. 1988;165:191–199. - PubMed
-
- De Francesco R, Behrens S E, Tomei L, Altamura S, Jiricny J. RNA-dependent RNA polymerase of hepatitis C virus. Methods Enzymol. 1996;275:58–67. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
