

Physical and functional interaction between the Y-box binding protein YB-1 and human polyomavirus JC virus large T antigen
- PMID: 10559330
- PMCID: PMC113067
- DOI: 10.1128/JVI.73.12.10146-10157.1999
Physical and functional interaction between the Y-box binding protein YB-1 and human polyomavirus JC virus large T antigen
Abstract
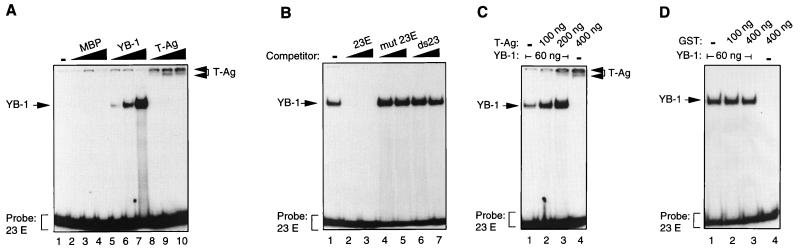
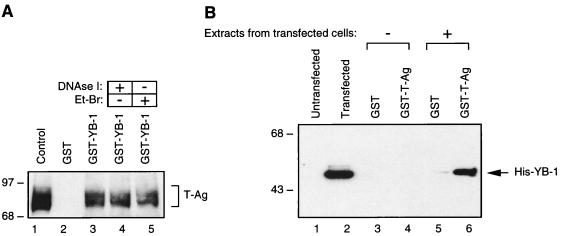
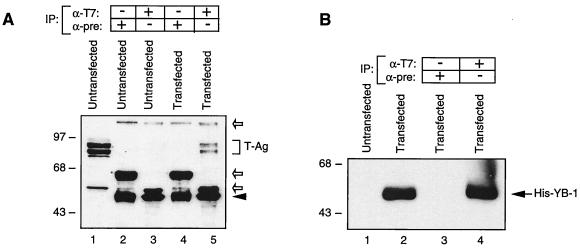
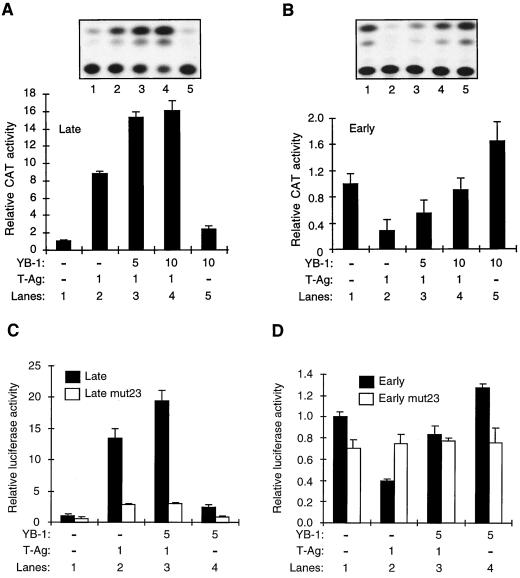
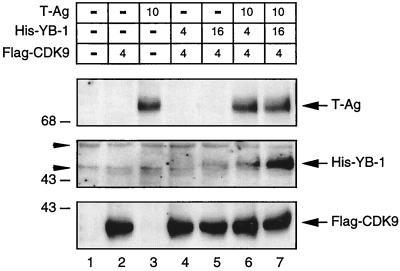
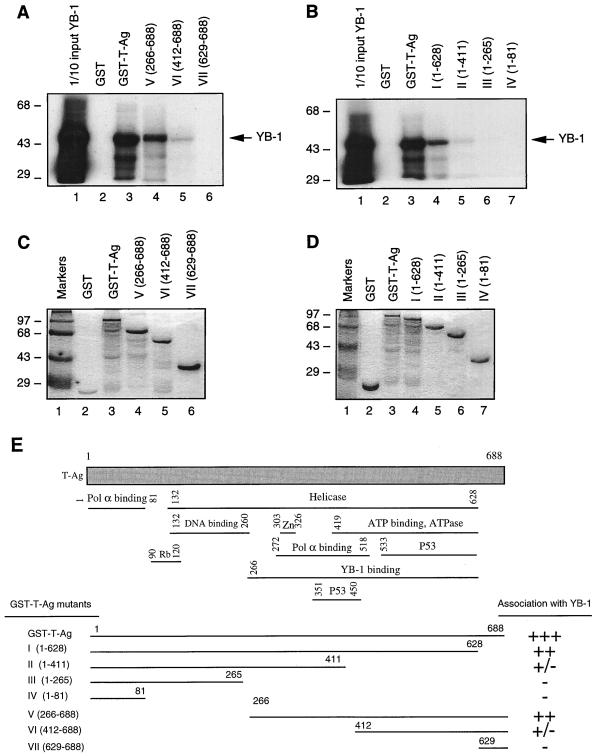
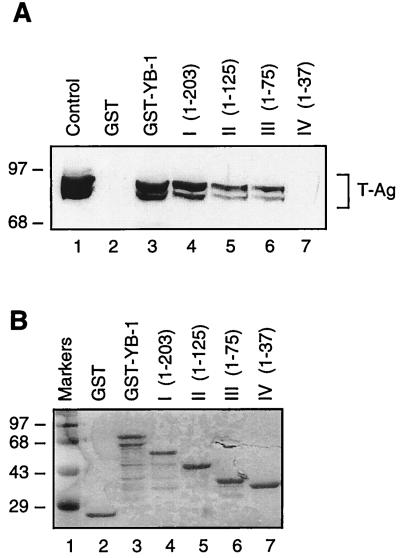
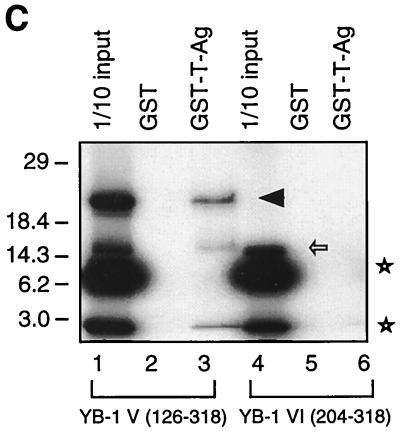
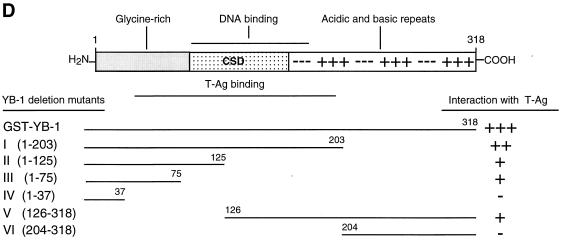
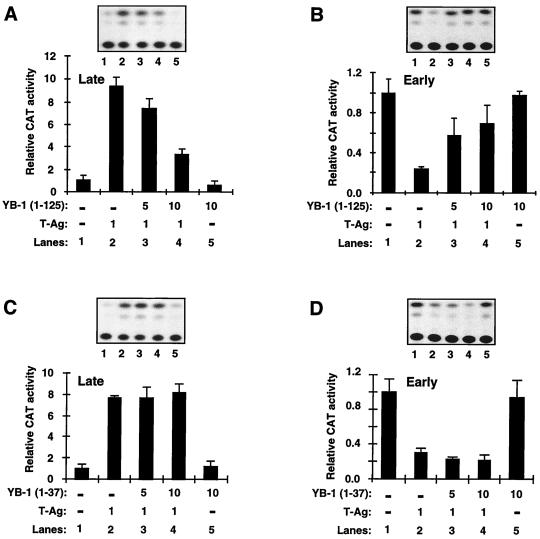
Y-box binding protein YB-1 is a member of a family of DNA and RNA binding proteins which have been shown to affect gene expression at both the transcriptional and translational levels. We have previously shown that YB-1 modulates transcription from the promoters of the ubiquitous human polyomavirus JC virus (JCV). Here we investigate the physical and functional interplay between YB-1 and the viral regulatory protein large T antigen (T-antigen), using JCV as a model system. Results of mobility band shift assays demonstrated that the efficiency of binding of YB-1 to a 23-bp single-stranded viral target sequence was significantly increased when T-antigen was included in the binding reaction mixture. Affinity chromatography and coimmunoprecipitation assays demonstrated that YB-1 and T-antigen physically interact with each other. Additionally, results of transcription studies demonstrated that these two proteins interact functionally on the JCV early and late gene promoters. Whereas ectopic expression of YB-1 and T-antigen results in synergistic transactivation of the viral late promoter, YB-1 alleviates T-antigen-mediated transcriptional suppression of the viral early promoter activity. Furthermore, we have localized, through the use of a series of deletion mutants, the sequences of these proteins which are important for their interaction. The T-antigen-interacting region of YB-1 is located in the cold shock domain of YB-1 and its immediate flanking sequences, and the YB-1-interacting domain of T-antigen maps to the carboxy-terminal half of T-antigen. Results of transient transfection assays with various YB-1 mutants and T-antigen expression constructs confirm the specificity of the functional interaction between YB-1 and T-antigen. Taken together, these data demonstrate that the cellular factor YB-1 and the viral regulatory protein T-antigen interact both physically and functionally and that this interaction modulates transcription from the JCV promoters.
Figures
References
-
- Ansari S A, Safak M, Gallia G L, Sawaya B E, Amini S, Khalili K. Interaction of YB-1 with human immunodeficiency virus type and TAR RNA modulates viral promoter activity. J Gen Virol. 1999;80:229–236. - PubMed
-
- Asakuno K, Kohno K, Uchiumi T, Kubo T, Sato S, Isono M, Kuwano M. Involvement of a DNA binding protein, MDR-NF1/YB-1, in human MDR1 gene expression by actinomycin D. Biochem Biophys Res Commun. 1994;199:1428–1435. - PubMed
-
- Bargou R C, Jurchott K, Wagener C, Bergmann S, Metzner S, Bommert K, Mapara M Y, Winzer K J, Dietel M, Dorken B, Royer H D. Nuclear localization and increased levels of transcription factor YB-1 in primary human breast cancers are associated with intrinsic MDR1 gene expression. Nat Med. 1997;3:447–450. - PubMed
-
- Berger J R, Concha M. Progressive multifocal leukoencephalopathy: the evolution of a disease once considered rare. J Neurovirol. 1995;1:5–18. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
