

Brome mosaic virus RNA replication proteins 1a and 2a colocalize and 1a independently localizes on the yeast endoplasmic reticulum
- PMID: 10559348
- PMCID: PMC113085
- DOI: 10.1128/JVI.73.12.10303-10309.1999
Brome mosaic virus RNA replication proteins 1a and 2a colocalize and 1a independently localizes on the yeast endoplasmic reticulum
Abstract
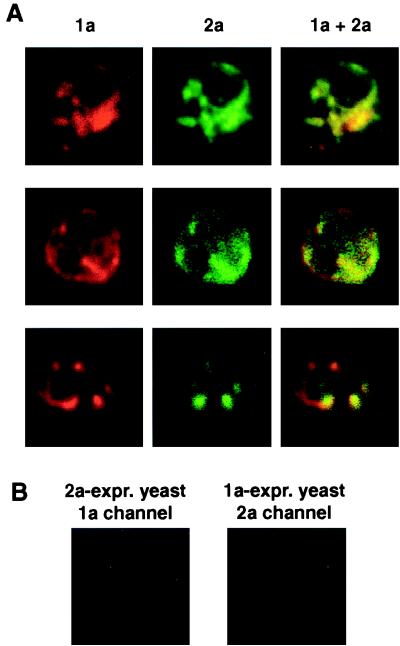
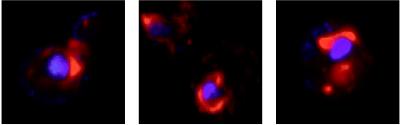
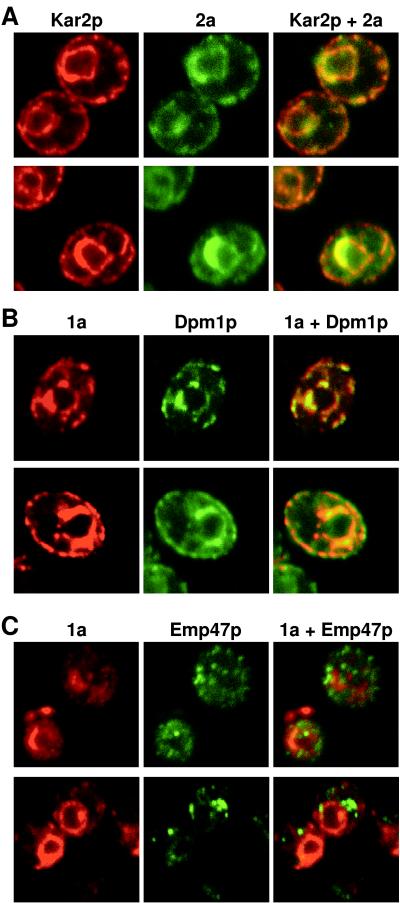
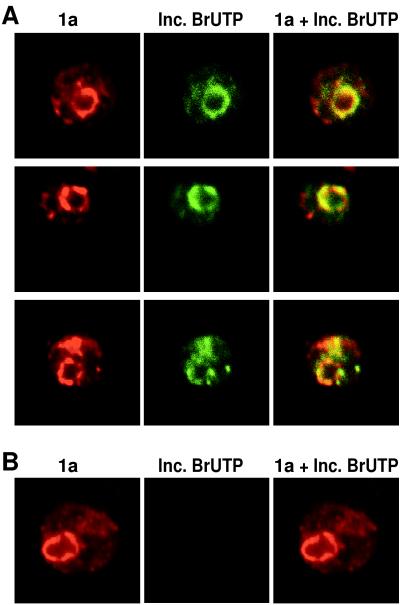
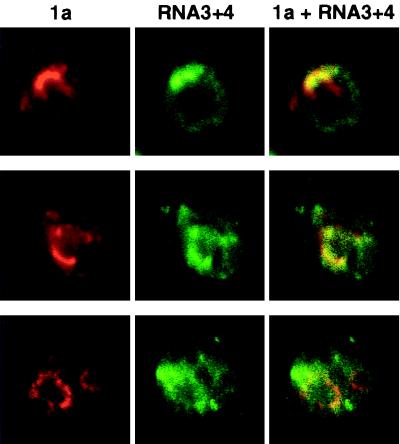
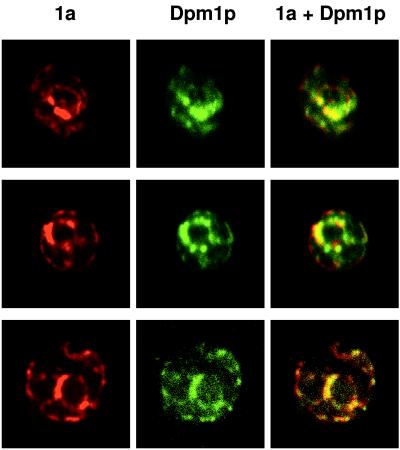
The universal membrane association of positive-strand RNA virus RNA replication complexes is implicated in their function, but the intracellular membranes used vary among viruses. Brome mosaic virus (BMV) encodes two mutually interacting RNA replication proteins: 1a, which contains RNA capping and helicase-like domains, and the polymerase-like 2a protein. In cells from the natural plant hosts of BMV, 1a and 2a colocalize on the endoplasmic reticulum (ER). 1a and 2a also direct BMV RNA replication and subgenomic mRNA synthesis in the yeast Saccharomyces cerevisiae, but whether the distribution of 1a, 2a, and active replication complexes in yeast duplicates that in plant cells has not been determined. For yeast expressing 1a and 2a and replicating BMV genomic RNA3, we used double-label confocal immunofluorescence to define the localization of 1a, 2a, and viral RNA and to explore the determinants of replication complex targeting. As in plant cells, 1a and 2a colocalized on and were retained on the yeast ER, with no detectable accumulation in the Golgi apparatus. 1a and 2a were distributed over most of the ER surface, with strongest accumulation on the perinuclear ER. In vivo labeling with bromo-UTP showed that the sites of 1a and 2a accumulation were the sites of nascent viral RNA synthesis. In situ hybridization showed that completed viral RNA products accumulated predominantly in the immediate vicinity of replication complexes but that some, possibly more mature cells also accumulated substantial viral RNA in the surrounding cytoplasm distal to replication complexes. Additionally, we find that 1a localizes to the ER when expressed in the absence of other viral factors. These results show that BMV RNA replication in yeast duplicates the normal localization of replication complexes, reveal the intracellular distribution of RNA replication products, and show that 1a is at least partly responsible for the ER localization and retention of the RNA replication complex.
Figures
References
-
- Ahlquist P, Luckow V, Kaesberg P. Complete nucleotide sequence of brome mosaic virus RNA3. J Mol Biol. 1981;153:23–38. - PubMed
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: John Wiley & Sons; 1987.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
