

In vivo regulation of hepatitis B virus replication by peroxisome proliferators
- PMID: 10559356
- PMCID: PMC113093
- DOI: 10.1128/JVI.73.12.10377-10386.1999
In vivo regulation of hepatitis B virus replication by peroxisome proliferators
Abstract
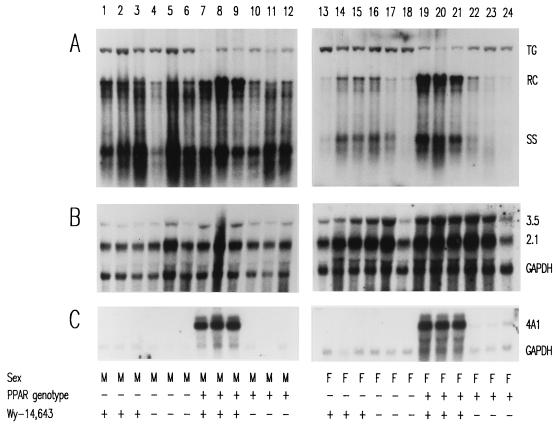
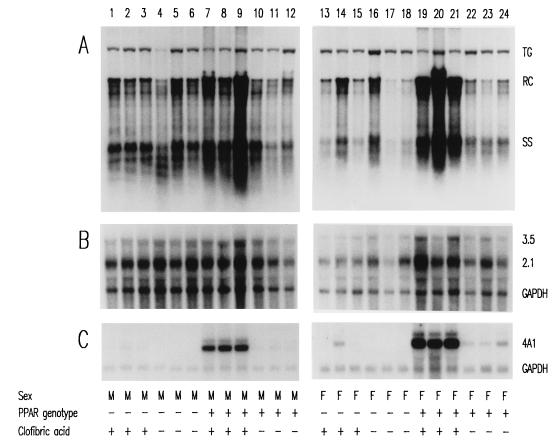
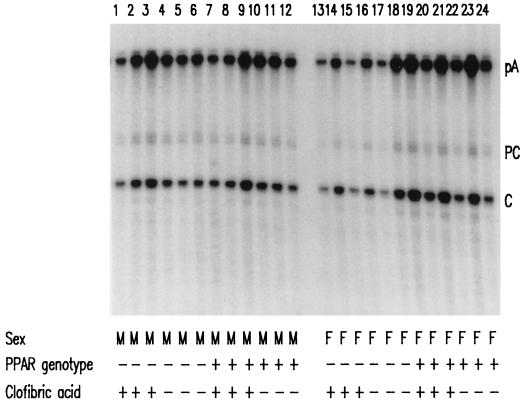
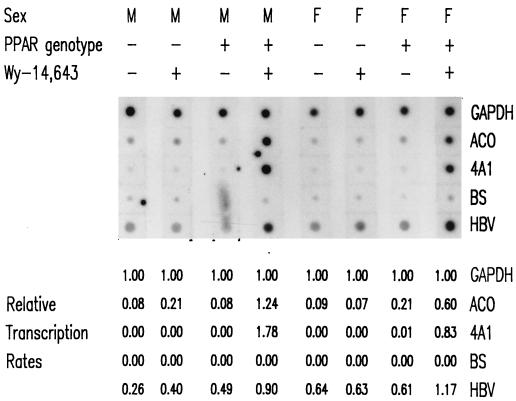
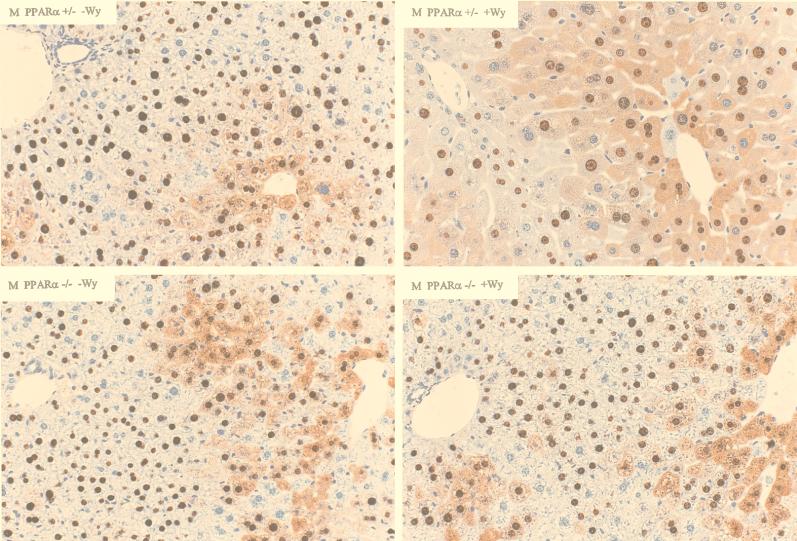
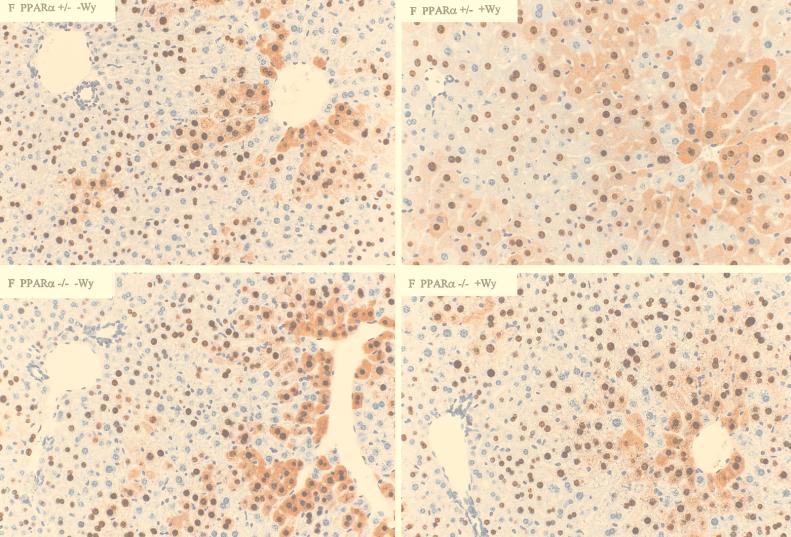
The role of the peroxisome proliferator-activated receptor alpha (PPARalpha) in regulating hepatitis B virus (HBV) transcription and replication in vivo was investigated in an HBV transgenic mouse model. Treatment of HBV transgenic mice with the peroxisome proliferators Wy-14,643 and clofibric acid resulted in a less than twofold increase in HBV transcription rates and steady-state levels of HBV RNAs in the livers of these mice. In male mice, this increase in transcription was associated with a 2- to 3-fold increase in replication intermediates, whereas in female mice it was associated with a 7- to 14-fold increase in replication intermediates. The observed increases in transcription and replication were dependent on PPARalpha. HBV transgenic mice lacking this nuclear hormone receptor showed similar levels of HBV transcripts and replication intermediates as untreated HBV transgenic mice expressing PPARalpha but failed to demonstrate alterations in either RNA or DNA synthesis in response to peroxisome proliferators. Therefore, it appears that very modest alterations in transcription can, under certain circumstances, result in relatively large increases in HBV replication in HBV transgenic mice.
Figures
References
-
- Block T M, Lu X Y, Mehta A, Park J, Blumberg B S, Dwek R. Role of glycan processing in hepatitis B virus envelope protein trafficking. Adv Exp Med Biol. 1998;435:207–216. - PubMed
-
- Buckwold V E, Chen M, Ou J H. Interaction of transcription factors RFX1 and MIBP1 with the gamma motif of the negative regulatory element of the hepatitis B virus core promoter. Virology. 1997;227:515–518. - PubMed
-
- Chen M, Ou J H. Cell type-dependent regulation of the activity of the negative regulatory element of the hepatitis B virus core promoter. Virology. 1995;214:198–206. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
